Bugün öğrendim ki: Coelacanthlar ve diğer lob yüzgeçli balıklar, diğer balıklara (örneğin köpekbalıkları, somon vb.) göre insanlara daha yakındır
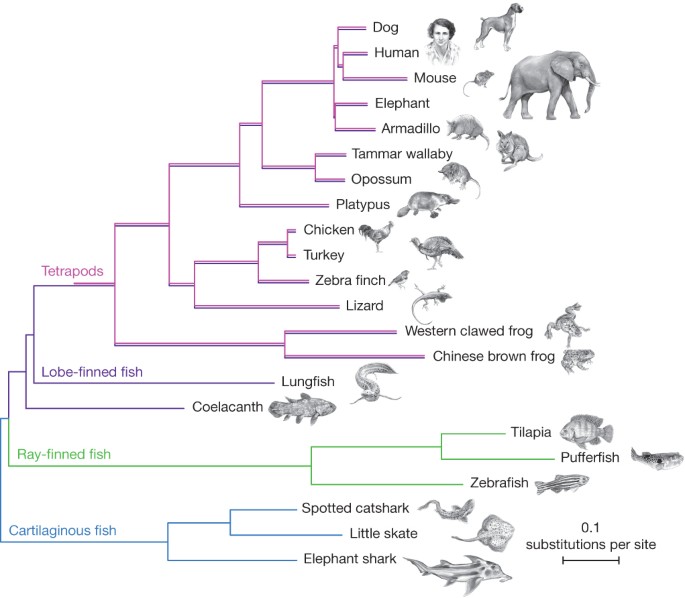
Modern koelakantın fosillerle olan morfolojik benzerliği, ona "yaşayan fosil"1 lakabını taktırmıştır. Bu durum, koelakant genomunun, dış görünümünün önerdiği gibi yavaş mı evrimleştiği sorusunu ortaya çıkarır. Daha önceki çalışmalar, Hox ve protokadherinler gibi birkaç gen ailesinin, diğer omurgalı soy hatlarına göre koelakantta protein kodlayan evrimin nispeten daha yavaş olduğunu göstermiştir8,10. Bu soruyu ele almak için, koelakant genomunun çeşitli özelliklerini diğer omurgalı genomlarıyla karşılaştırdık.
Protein kodlayan gen evrimi, yukarıda açıklanan filogenomik veri kümesi (251 birleştirilmiş protein) kullanılarak incelendi (Şekil 1). Taksonlar arasındaki karşılıklı mesafeler, ağaç dalları uzunluklarından, ortalama ikame oranlarının eşitliğini test etmek için daha önce önerilmiş iki küme testini kullanarak hesaplandı20. Ardından, aşağıdaki türler ve tür kümeleri (koelakant, akciğerli balık, tavuk ve memeliler) için, her birinin, üç kıkırdaklı balıktan (fil balığı, küçük paten ve benekli kedi köpekbalığı) oluşan bir dış gruba göre ortalama uzaklıklarını belirledik. Son olarak, her bir tür ve tür kümesi çiftinde, kıkırdaklı balık dış grubuna olan uzaklıkta önemli bir fark olup olmadığını, Z istatistiği kullanarak test ettik. Kıkırdaklı balık dış grubuna olan bu mesafeler karşılaştırıldığında, test edilen koelakant proteinlerinin, akciğerli balıktan (söz konusu yerde 1.05 ikame), tavuktan (söz konusu yerde 1.09 ikame) ve memeliden (söz konusu yerde 1.21 ikame) ortoloğlardan (tüm durumlarda P < 10−6) önemli ölçüde daha yavaş evrimleştiğini (söz konusu yerde 0.890 ikame) bulduk (Ek Veri 5). Ayrıca, Şekil 1'de görülebileceği gibi, koelakanttaki ikame oranı, iki soy hattı ayrıldıktan beri dört ayaklılarda yaklaşık olarak yarıya indi. Bir Tajima'nın göreli oran testi21, koelakantın önemli ölçüde daha yavaş protein evrimi oranını doğruladı (P < 10−20) (Ek Veri 6).
Ardından, koelakant genomundaki transpoze edilebilir elementlerin bolluğunu inceledik. Teorik olarak, transpoze edilebilir elementler, yeni düzenleyici elementler ve ekzonlar oluşturmak için exaptation şablonları oluşturarak ve genomik yeniden düzenlemenin substratları olarak görev yaparak bir türün evrimine en büyük katkılarını sağlayabilir22. Koelakant genomunun, çeşitli transpoze edilebilir element üst ailelerini içerdiğini ve nispeten yüksek bir transpoze edilebilir element içeriğine (yüzde 25) sahip olduğunu bulduk; bu sayı muhtemelen bir taslak montaj olduğu için bir küçümsemedir (Ek Not 5 ve Ek Tablolar 7–10). RNA-seq verilerinin ve bireysel transpoze edilebilir element kopyalarının konsensüs dizilerinden ayrışmasının analizi, 14 koelakant transpoze edilebilir element üst ailesinin şu anda aktif olduğunu göstermektedir (Ek Not 6, Ek Tablo 10 ve Ek Şekil 5). Koelakant genomunun, şu anda birçok diğer genomla benzer şekilde, transpoze edilebilir elementlerin hem bolluğunu hem de aktivitesini gösterdiğini sonucuna varıyoruz. Bu, gözlemlenen yavaş protein evriminin aksine bir durumdur.
Koelakant genomunda ve dört ayaklı genomlarındaki kromozom kırılma noktalarının analizleri, sinyenin geniş bir şekilde korunmasını ortaya koymakta ve büyük ölçekli yeniden düzenlemelerin koelakant soy hattında genellikle düşük bir hızda meydana geldiğini göstermektedir. Bu yeniden düzenleme sınıflarının analizleri, daha önce yayınlanmış dört ayaklı soy hatlarında meydana geldiği bilinen birkaç fisyon olayını23 ve koelakant soy hattında veya erken dört ayaklı soy hattında meydana gelen en az 31 interkromozomal yeniden düzenlemeyi (1 milyon yılda 0.063 füzyon) tespit etti; bu, semender soy hattındaki 20 olayla (1 milyon yılda 0.054 füzyon) ve Xenopus soy hattındaki 21 olayla (1 milyon yılda 0.057 füzyon) karşılaştırılabilir23 (Ek Not 7 ve Ek Şekil 6). Genel olarak, bu analizler, koelakant soy hattındaki karyotipik evrimin, memeli olmayan dört ayaklılar24'ünkiyle benzer bir şekilde nispeten yavaş bir hızda gerçekleştiğini göstermektedir.
Ayrı bir analizde, sırasıyla Afrika ve Endonezya sularında bulunan iki koelakant türü olan L. chalumnae ve L. menadoensis arasındaki evrimsel ayrışmayı da inceledik. Mitokondriyal DNA'nın önceki analizi, yüzde 96'lık bir dizi özdeşliği gösterdi, ancak tahmini ayrışma zamanları 6 ila 40 milyon yıl arasında geniş bir yelpazede değişiyor25,26. L. menadoensis'in karaciğer ve testis transkriptomik verilerini27 L. chalumnae genomuyla karşılaştırdığımızda, yüzde 99.73'lük bir özdeşlik bulduk (Ek Not 8 ve Ek Şekil 7), oysa 20 dizilenmiş L. menadoensis bakteriyel yapay kromozom (BAC) ile L. chalumnae genomu arasındaki hizalamalar yüzde 98.7'lik bir özdeşlik gösterdi (Ek Tablo 11 ve Ek Şekil 8). Hem genik hem de genomik ayrışma oranları, insan ve şempanze genomları arasında görülenlerle (sırasıyla yüzde 99.5 ve yüzde 98.8; 6 ila 8 milyon yıl önceki ayrışma zamanı)28 benzerdir, oysa Latimeria'daki moleküler evrim oranları, koelakantta görülen daha yavaş ikame oranı da dahil olmak üzere çeşitli faktörlerden etkilenmektedir. Bu, iki koelakant türü için biraz daha uzun bir ayrışma zamanını düşündürmektedir.
Genom dizisi oluşturulmuş en yakın sucul atasımıza ait tür olarak, koelakant, omurgalıların karasal ortama başarılı bir şekilde uyum sağlamasıyla ilişkili genomik değişiklikleri belirlemek için benzersiz bir fırsat sunmaktadır.
Omurgalıların karada yaşadığı 400 milyon yıldan fazla bir süre boyunca, yeni ortamlarında var olmak için gerekli olmayan bazı genler ortadan kaldırılmıştır. Su-kara geçişinin bu yönünü anlamak için, Latimeria genomu anotasyonlarını tarayarak, tüm kemikli balıkların (koelakant dahil) son ortak atasında bulunan, ancak dört ayaklı genomlarında bulunmayan genleri belirledik. Koelakant verilerine dayanarak, 50'den fazla genin, fibroblast büyüme faktörü (FGF) sinyallemesi, TGF-β ve kemik morfogenetik protein (BMP) sinyallemesi ve WNT sinyalleme yollarının bileşenlerini ve ayrıca birçok transkripsiyon faktörü genini de içeren, kaybolduğu çıkarıldı (Ek Veri 7 ve Ek Şekil 9). Bu geçişte kaybolan genlerin önceki çalışmaları, sadece teleost balıklarını dört ayaklılarla karşılaştırabilirdi, bu da gen içeriğindeki farklılıkların, dört ayaklı veya loblu yüzgeçli balık soy hatlarında kayıp nedeniyle olabileceği anlamına geliyordu. Daha önce dört ayaklılarda bulunmadığı gösterilen dört genin (And1 ve And2 (kaynak 29), Fgf24 (kaynak 30) ve Asip2 (kaynak 31)), Latimeria'da gerçekten mevcut ve sağlam olduğunu doğrulayabildik; bu da, dört ayaklı soy hattında kayboldukları fikrini desteklemektedir.
Zebra balığı verilerini kullanarak, dört ayaklılarda kaybolan 50'den fazla geni işlevsel olarak anotladık (gen ifadesi, nakavt ve nakavtlar). Birçok gen, önemli gelişimsel kategorilere sınıflandırıldı (Ek Veri 7): yüzgeç gelişimi (13 gen); otolit ve kulak gelişimi (8 gen); böbrek gelişimi (7 gen); gövde, somite ve kuyruk gelişimi (11 gen); göz (13 gen); ve beyin gelişimi (23 gen). Bu, sudan karaya geçişteki morfolojik geçişteki kritik karakterlerin (örneğin, yüzgeçten uzuva geçiş ve kulağın yeniden modellenmesi), dört ayaklılara yol açan filogenetik dal boyunca belirli genlerin kaybında yansıtıldığını ima eder. Bununla birlikte, bir organizmanın temel vücut planının gelişiminden sorumlu olan homeobox genleri, Latimeria, ışın yüzgeçli balıklar ve dört ayaklılar arasında sadece hafif farklılıklar gösterir; bu gen ailesinin protein kodlayan kısmının, diğer birkaç genin (Ek Not 9, Ek Tablolar 12–16 ve Ek Şekil 10) yanı sıra, omurgalı karasal geçişi sırasında büyük ölçüde korunmuş göründüğü söylenebilir (Ek Şekil 11).
Omurgalıların yeni bir karasal ortama geçişiyle birlikte, gen içeriğinde değil, aynı zamanda mevcut genlerin düzenlenmesinde de değişiklikler meydana geldi. Korunmuş kodlanmayan elementler (CNE'ler), gen düzenleyici elementler için güçlü adaylardır. Bunlar, promotörler, güçlendiriciler, bastırıcılar ve yalıtıcılar olarak görev yapabilir32,33 ve evrimsel değişimin ana kolaylaştırıcıları olarak kabul edilir34. Dört ayaklıların en son ortak atasında ortaya çıkan CNE'leri belirlemek için, çeşitli kemikli omurgalı (yani, ışın yüzgeçli balık, koelakant ve dört ayaklı) soy hatlarında evrimleşen CNE'leri tahmin ettik ve bunları olası dal noktalarına atadık. CNE'leri tespit etmek için, insan genomunda korunmuş diziler, MULTIZ hizalamalarını kullanarak kemikli omurgalı genomlarında tanımlandı ve ardından bilinen protein kodlayan diziler, çevrilmemiş bölgeler (UTR'ler) ve bilinen RNA genleri hariç tutuldu. Analizimiz, koelakant soy hattının ayrışmasından sonra ortaya çıkan 44.200 atasal dört ayaklı CNE'si belirledi. Bunlar, kemikli omurgalı soy hattında kısıtlama altında bulunan 739.597 CNE'nin yüzde 6'sını temsil ediyor. Atasal dört ayaklı CNE'leri, bir transkripsiyonel koaktivatör olan p300'e karşı antikorlar kullanılarak elde edilen fare embriyosu ChIP-seq (kromatin bağışıklık çökeltmesi ardından dizileme) verileriyle karşılaştırdık. Bu, aday CNE'lerimiz için p300 bağlama bölgelerinde yedi kat zenginleşmeyle sonuçlandı ve bu CNE'lerin gerçekten gen düzenleyici elementler açısından zengin olduğunu doğruladı.
Her dört ayaklı CNE'si, transkripsiyon başlangıç noktası en yakın olan gene atandı ve bu genler için gen ontolojisi kategorisi zenginleştirmesi hesaplandı. En zengin kategoriler, koku algısıyla (örneğin, kokunun duyusal algısı, kimyasal uyaranın algılanması ve koku alma reseptör aktivitesi) ilgiliydi. Bu, teleostlarla karşılaştırıldığında dört ayaklılarda koku alma reseptör ailesi genlerinin dikkate değer bir şekilde genişlemesiyle uyumludur ve karasal yaşam tarzının bir parçası olarak, havadaki koku maddelerini algılamak için daha sıkı düzenlenmiş, daha büyük ve daha çeşitli bir koku alma reseptör repertuarının gerekliliğini yansıtabilir. Diğer önemli kategoriler arasında, karasal geçiş için gerekli vücut planı değişiklikleriyle uyumlu olan morfogenez (radyal desen oluşumu, arka bacak morfogenezi, böbrek morfogenezi) ve hücre farklılaşması (endotel hücre kaderi taahhüdü, epitel hücre kaderi taahhüdü) ve ayrıca omurgalıların karada karşılaşacağı yeni patojenleri ele almak için varsayılan cevap farklılıklarını yansıtan immünoküre VDJ rekombinasyonu bulunur (Ek Not 10 ve Ek Tablolar 17–24).
Dört ayaklıların önemli bir yeniliği, parmaklarla karakterize edilen uzuvların evrimidir. Uzuv iskeleti, bir stilopoddan (humerus veya femur), bir zeugopoddan (radius ve ulna, veya tibia ve fibula) ve bir autopoddan (bilek veya ayak bileği ve parmaklar) oluşur. Autopod'un kökenleri hakkında iki ana hipotez vardır; bunun dört ayaklıların yeni bir özelliği olduğu ve balıkların yüzgeçlerinde atalarının olduğu35 (Ek Not 11 ve Ek Şekil 12). Burada, bu hipotezleri ele almak için ışın yüzgeçli balıklar, koelakant ve dört ayaklılarda uzuv gelişimi üzerindeki Hox düzenlemesini inceliyoruz.
Farede, geç evre parmak güçlendiricileri, HOX-D kümesine yakın bir gen çölünde bulunur36. Burada, koelakantın HOX-D santromerik gen çölünün, dört ayaklılar ve ışın yüzgeçli balıklarınkiyle hizalamasını sağlıyoruz (Şekil 2a). Daha önce bu gen çölünde tanımlanan altı cis-düzenleyici diziden36, üç dizinin, sadece dört ayaklılarda dizi korunumu gösterdiği görülmektedir (Ek Şekil 13). Bununla birlikte, bir düzenleyici dizi (ada 1), dört ayaklılar ve koelakant tarafından paylaşılır, ancak ışın yüzgeçli balıklar tarafından paylaşılmaz (Şekil 2b ve Ek Şekil 14). Farede geçici bir transgenik deneyde test edildiğinde, ada 1'in koelakant dizisi, bir uzuv spesifik modelinde rapor geninin ifadesini yönlendirebildi (Şekil 2c). Bu, ada 1'in, dört ayaklıların balık atasında loblu yüzgeç gelişimi güçlendiricisi olduğunu ve daha sonra modern dört ayaklıların autopod güçlendiricisine dahil edildiğini düşündürmektedir. Bu durumda, autopod gelişimi düzenlemesi, atasal loblu yüzgeçli balık düzenleyici bir elementten türetilmiştir.
Üre döngüsündeki değişiklikler, karaya geçişle ilişkili adaptasyonların aydınlatıcı bir örneğini sunmaktadır. Azotun atılımı, karasal omurgalılar için önemli bir fizyolojik zorluktur. Sulu ortamlarda, birincil azotlu atık ürünü, çevredeki su tarafından toksik seviyelere ulaşmadan önce kolayca seyreltilen amonyaktır, ancak karada, bunun yerine üre veya ürik asit gibi daha az toksik maddeler üretilmelidir (Ek Şekil 15). Amfibilerde, bazı kaplumbağalarda ve memelilerde yaygın ve neredeyse sadece üre atılımının görülmesi, ürenin ana azotlu atık ürünü olarak kullanılmasının, omurgalıların sudan karaya geçişindeki temel bir yenilik olduğu hipotezine yol açmıştır37.
Koelakant ve akciğerli balıktan gen dizilerinin kullanılabilir hale gelmesiyle, bu hipotezi test etmek mümkün hale geldi. HYPHY paketinde38, çoklu tür dizisi hizalaması boyunca farklı dallarda ve farklı bölgelerde (kodonlar) sinonim (dS) ile sinonim olmayan (dN) ikame oranlarının (ω değerleri) oranını tahmin eden bir dal-siten modeli kullandık. Hepatik üre döngüsünün hız sınırlayıcı enzimi olan karbamil fosfat sentaz I (CPS1) için, sadece ağacın bir dalında güçlü bir seçilim imzası gözlemlendi (P = 0.02), yani dört ayaklılara giden dal ve amniyotlara giden dal (Şekil 3); bu döngüdeki diğer enzimlerde bir seçilim imzası gözlemlenmedi. Tersine, mitokondriyal arginaz (ARG2), arginin metabolizmasının bir yan ürünü olarak ektrahepatik üre üretir, ancak azotlu atık bertarafı için üre üretiminde yer almaz, omurgalılarda herhangi bir seçilim kanıtı göstermedi (Ek Şekil 16). Bu, omurgalıların karasal geçişi sırasında hepatik üre döngüsünde uyarlamalı evrimin gerçekleştiği sonucuna varmamızı sağlar. Ek olarak, koelakant ve dört ayaklılar arasında değişen CPS1'in beş amino asitinden üçünün önemli alanlarda (iki ATP bağlama bölgesi ve alt birim etkileşim alanı) ve dördünün insan hastalarında mutasyona uğradığında bir arızalı enzime neden olduğu bilinmektedir39.
Karasal yaşam tarzına uyum, gelişen embriyonun ve fetüsün fizyolojik ortamında önemli değişiklikler gerektirdi ve amniyot memelilerin ekstraembriyonik zarlarının evrimini ve özelleşmesini sağladı40. Özellikle plasenta, anne ve fetüs arasında gaz ve besin alışverişinde bulunmak için kritik önem taşıyan karmaşık bir yapıdır ve aynı zamanda önemli bir hematopoez yeridir41.
Dört ayaklılarda, özelikle eutiryan plasenta dahil olmak üzere ekstraembriyonik yapıların evriminde rol oynamış olabilecek, koelakant HOX-A kümesinin bir bölgesini tanımladık. Koelakant Hoxa14-Hoxa13 bölgesinin, boynuz köpekbalığı, tavuk, insan ve farenin homolog bölgeleriyle küresel hizalaması, koelakant Hoxa14 geninin hemen yukarısında bir CNE ortaya koydu (Ek Şekil 17a). Bu korunmuş kısım, teleost balıklarda bulunmaz, ancak tavuğun, insanın ve farenin Hoxa14 ortologlarına sahip olmamasına ve boynuz köpekbalığı Hoxa14 geninin sözde bir gen haline gelmesine rağmen, boynuz köpekbalığı, tavuk, insan ve fare arasında oldukça korunmuştur. Bu CNE, HA14E1, Latimeria'daki Hoxa14 geninin proksimal promotör-güçlendirici bölgesine karşılık gelir. HA14E1, fare, insan ve diğer tüm dizilenmiş memeliler arasında yüzde 99'dan fazla özdeşdir ve bu nedenle aşırı korunmuş bir element olarak kabul edilecektir42. Yüksek düzeyde korunma, promotör aktivitesine sahip olan bu elementin, amniyotlarda Hoxa14 geninin kaybına rağmen diğer işlevler için devşirilmiş olabileceğini düşündürmektedir (Ek Şekil 17bc). Farede geçici bir transgenik deneyde insan HA14E1'in ifadesi, gün 11.5'te embriyo propriosunda önemli bir ifade vermedi (bilgi, VISTA güçlendirici tarayıcı web sitesinde çevrimiçi olarak mevcuttur; http://enhancer.lbl.gov/cgi-bin/imagedb3.pl?form=presentation&show=1&experiment_id=501&organism_id=1), bu da konumunun aksiyel yapıları kaudalda düzenleyeceğini tahmin etmesi nedeniyle beklenmedik bir durumdu43. Tavuk embriyolarında, tavuk HA14E1'i kullanarak yapılan benzer bir deney, anteroposterior eksende de herhangi bir aktivite göstermedi. Bununla birlikte, tavuk embriyosunun ekstraembriyonik alan vasculosasında güçlü bir ifade gözlemlendi (Şekil 4a). Fare embriyolarında bir Latimeria BAC Hoxa14-rapor transgeninin incelenmesi, Hoxa14 geninin embriyonal gün 8.5'te ekstraembriyonik bir bölgede hücrelerin bir alt kümesinde özel olarak ifade edildiğini gösterdi (Şekil 4b).
Bu bulgular, HA14E1 bölgesinin, farenin allantoisinde ifade edildiği bilinen ve memeli plasentasının erken oluşumu için kritik olan posterior HOX-A genlerinin (Hoxa13, Hoxa11 ve Hoxa10) düzenlemesini koordine etmek için evrimsel olarak işe alınmış olabileceğini düşündürmektedir. Latimeria'nın bir plasentası olmasa da, canlı yavru doğurur ve çok büyük, vaskülerleşmiş yumurtalara sahiptir, ancak Hoxa14, HA14E1 güçlendiricisi ve koelakanttaki kan adacığı oluşumu arasındaki ilişki bilinmemektedir.