Bugün öğrendim ki: böcekler aslında ışığa çekilmezler ancak Dorsal Işık Tepkisi adı verilen yerleşik bir refleks nedeniyle ışığı sırtlarının üstünde tutmaya çalışırlar. Bu, sırtlarını gökyüzüyle karıştırarak ışığa doğru çevirmelerine neden olur ve bu da yapay ışık kaynaklarının etrafında dönmelerine neden olur
Yapay nokta ışık kaynağı böceklerde anormal uçuş davranışına neden olur
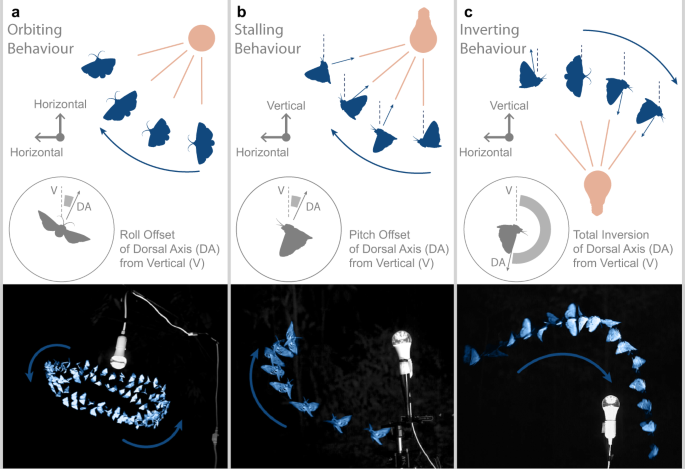
477 stereo-videografik alan kaydı (ortanca süre 1,7 sn IQR 1,9 sn) (Ek Şekil 1 ve Ek Veri 1, 2) boyunca, yapay ışığın yakınında uçan 10 böcek takımında gözlemlenen üç görsel olarak belirgin davranışsal motifini (Tablo 1) belirledik (Tablo 2). Yörünge uçuşu, sürekli hızla ışık etrafındaki nispeten sabit dairesel uçuş yoluyla tanımlanabilir (Şekil 1a ve Ek Video 1). Böcek, vücudu ışığa doğru yanal olarak eğik (eğimli veya yuvarlanmış) bir şekilde sabit bir eğimli duruşu koruduğu görülmüştür. Yörünge uçuş, düşük rüzgar koşullarında (<1 m/s) yaygındı ve rüzgar estiğinde böcekler dağılıyordu. Durgunluk, böceğin ışık kaynağından uzağa bakarken dik bir tırmanışla karakterize edildi (Şekil 1b ve Ek Video 2), böcek ilerlemeyi bırakana kadar hızını kaybetti. Böcek, bir ışık kaynağının hemen üstünden uçtuğunda (Şekil 1c ve Ek Video 3) böceğin duruşunun ters çevrilmesi (yuvarlanma veya eğim yoluyla) yere dik bir dalışa neden oldu. Bu motifleri tüm koşullar altında ışık kaynağının etrafında uçan böceklerde gözlemledik, ancak karanlıkta uçan böceklerin videolarında nadiren (%2'den az), bu nedenle bunları anormal olarak nitelendirdik (Tablo 1). Işığın altına düştükten sonra, böcekler sık sık kendilerini düzelttiler, ancak yalnızca ışığın üzerine tırmandılar ve bir kez daha ters döndüler. Bu uçuşlar sırasında, böcekler sürekli olarak sırt eksenlerini ışık kaynağına doğru yönlendirdiler, bu durum sürekli uçuşu engellemiş ve düşmeye yol açmış olsa bile.
Hareket yakalama, ışığa doğru sırt eğimini nicel olarak ölçer
Alan kayıtlarımızda gözlemlenen davranışı nicel olarak anlamak için, kontrollü bir davranışsal alanda uçan böcekleri kaydetmek için böcek ölçeğinde hareket yakalama kullandık (ortanca izleme süresi 1,7 sn IQR 4,3 sn) (Ek Şekil 2 ve Ek Veri 3). Hareket yakalama sistemimiz, görüşlerini bozmadan böceklerin toraksına monte edilmiş özel bir işaret çerçevesini (<%5 böcek vücut ağırlığı) izlemek için kızılötesi ışık kullandı. L şeklinde düzenlenmiş üç işaret, uzayda çerçeve dönmelerini ve ötelemelerini ve dolayısıyla böceğin sırt eksenini ölçmemize olanak sağladı35. Bu deney için, sistemi kabul edilebilir doğrulukla kayıt hacmi için ayarladık (işaret kalıntıları <0,24 mm veya en küçük böcek için <7° mutlak oryantasyon hatası).
İki metre çapında silindirik bir çadır içinde üç farklı ışık kaynağının etrafında farklı böcek türlerini uçurduk: bir UV LED ampul (395 nm), bir UV-Mavi Aktinik tüp (spektral tepe noktası 370 nm) ve soğuk beyaz bir LED ampul, böcek için başka hiçbir ışık kaynağı belirgin değildi. Genellikle ışık tuzağına yakalanmasıyla ilişkilendirilmeyen gündüz türlerini test etmek için, Yaygın Okçu (Sympetrum striolatum) (n = 12) ve Göçmen Okçu (Aeshna mixta) (n = 2) yusufçuklarını kullandık. Gece türleri için, Sarı Alt Kanatlı Güveleri (Noctua pronuba ve Noctua fimbriata) (sırasıyla n = 8 ve n = 2) ve Lorquin Atlas Güvesi (Attacus lorquinii) (n = 3) kullandık. Bu dört türde 538 sürekli uçuş yörüngesi kaydettik (Ek Veri 4, Tablo 3).
Bir nokta ışık kaynağının etrafında uçarken, uçuşlar, alanda gözlemlenen motiflerle açıklanan şekilde açıkça bozuldu (Ek Video 4). Üstten bakıldığında uçuş yörüngeleri (Şekil 2a), ışığa doğru çok az doğrudan uçuşla ışığın etrafında yörünge uçuşunu gösterir. 4 türün hız vektörlerini zemin düzlemine yansıttık ve bunları ışığın anlık yönüyle karşılaştırdık (Şekil 2b). Her dört durumda da, hız vektörü ışığın yönünden dik olarak kuvvetlice yoğunlaştı ve ışığa doğru doğrudan uçma fikrini çürütüyor. Rayleigh z-testi kullanarak, hız vektörlerinin kümelenmesinin tüm türler için anlamlı olduğunu doğrulamak için test ettik (S. striolatum, z = 1356.10, p < 0.001, n = 7032 alt örneklenmiş yörünge noktası; A. mixta, z = 102.09, p < 0.001, n = 1085; Noctua sp., z = 624.90, p < 0.001, n = 1759, A. lorquiini, z = 359.30, p < 0.001, n = 1403). Uçuş alanında, Yaygın Okçu yusufçukları geniş spektrumlu yayılmış gölgelik ışığı altında (z = 0,43, p = 0,36, n = 269) veya zifiri karanlıkta (z = 0,43, p = 0,65, n = 304) böyle bir yörünge uçuş deseni göstermez. Bu, Yörünge uçuşunun muhafazaya değil, UV ışık kaynağına bağlı olduğunu gösterir (Şekil 2).
İşaret çerçevesi verileri, böceklerin sırtlarını ışık kaynağına doğru kuvvetlice eğdiğini ortaya koydu (Şekil 3). Zemin düzlemine yansıtılan sırt eksenini incelemek, vücut eğiminin dört türde de ışık yönüyle 1:1 oranında kuvvetlice eşleştiğini gösterdi (Şekil 3a). Böcekler, etrafında uçarken sırt eksenlerini doğrudan ışığa doğru eğdiler. Normalize edilmiş yansıtılan sırt eksenleri ile ışık kaynağı yönü arasındaki nokta çarpımını bir indeks olarak aldık: -1 (ışık kaynağından uzağa) ile 1 (ışık kaynağına doğru) arasında değişmektedir. Bir ışık kaynağının etrafında uçan böcekler için indeks değerlerimiz S. striolatum için 0,84 (n = 9904 kare), A. mixta için 0,79 (n = 1416), Noctua sp. için 0,82 (n = 1563) ve A. lorquinii için 0,82 (n = 1357) idi, bu da her türde ışığa doğru güçlü bir sırt eğimini göstermektedir. Aksine, ışık kapalıyken S. striolatum'un eğim yönü indeksi 0,17 (n = 713) idi ve bu da karanlıkta zayıf bir sırt eğim tutarlılığını göstermektedir.
Dört türün ışık bozulmasına uğramış uçuş duruş dağılımını, eğim ve açısal yön kompozisyonlarını çizerek daha ayrıntılı olarak inceledik (Şekil 3b). Bunu bozulmamış uçuşlarla karşılaştırmak için, yusufçukların arenayı tavandan aydınlatan parlak, geniş spektrumlu lambalar altında uçmalarına izin verdik. İki güve türü için, genel ışık seviyelerini düşük tutarken, difüz bir UV-Mavi tavan oluşturmak için tek bir aktinik tüp kullandık. Kontrol koşulları altında, dört tür de dönüş, tırmanma veya iniş gibi rutin manevralar gerektiğinde, vücut eğim açısı 30°'nin altında olmak üzere (eğim açısı ortalamaları ± çeyrek aralık: S. striolatum için 11,9 ± 14,8°, Noctua sp. için 13,9° ± 22,7°, A. mixta için 20,2 ± 21,4° ve A. lorquinii için 8,7° ± 11,2°) tipik seyir seviyesi uçuş dağılımı gösterdi. Bir nokta ışık kaynağının yakınındaki tüm türlerin eğim-açılma dağılımları, kontrollerinden önemli ölçüde farklıydı. S. striolatum ve Noctua sp., nokta kaynaklarının yakınında güçlü ve yüksek bir eğim gösterdi. Daha büyük A. mixta ve A. lorquinii daha az tutarlı bir vücut duruşu gösterdi, ancak bir nokta ışık kaynağının yakınında eğim-açılma dağılımlarını daha yüksek değerlere kaydırdı. Tüm türler bir nokta ışık kaynağının yakınında bir miktar daha yüksek eğim gösterdi (eğim açısı ortalamaları ± çeyrek aralık: S. striolatum için 43,8 ± 39,2°, Noctua sp. için 48,0° ± 30,7°, A. mixta için 29,3 ± 30,8° ve A. lorquinii için 30,7° ± 31,8°) (Wilcoxon rank sum; S. striolatum, z = 19,91, p < 0,001, Noctua sp., z = 11,18, p < 0,001, A. mixta, z = 4,32, p < 0,001, A. lorquiini, z = 16,85.91, p < 0,001). Bu veriler, böcekler sırt eksenlerini ışığa hizalamaya çalışırken, bir nokta ışık kaynağının duruş kontrolünü önemli ölçüde değiştirdiğini göstermektedir.
Gök benzeri yapay ışık normal uçuşu geri yükler
Böcekleri ışık tuzağına yakalamanın bilinen bir yöntemi, parlak bir ışığı beyaz bir çarşafa tutmaktır36. Alanda, aşağıya doğru bakan (ampul yukarıda gizli) beyaz bir kumaş çarşaf üzerine gizlenmiş bir UV ışık kaynağını filme aldık. Bu kayıtlar sırasında, böceklerin yere çakılmadan önce havada ters döndüğünü ve takla attığını gözlemledik (Şekil 4a). Bu tuzaklama etkisi DLR tarafından aracılık ediliyorsa, böceklerin doğal ipuçlarıyla eşleşen aksi takdirde benzer ışık kaynakları tarafından yakalanmasını beklemiyoruz. Aynı gizlenmiş UV ampulü yukarıya doğru, yukarıya gerilmiş beyaz bir çarşafa tuttuğumuzda, UV-Mavi ışığın gökyüzüne benzer şekilde difüz bir gölgelik olarak aşağı yansıdığı bir koridor yarattı. Bu düzenlemede, böcekler ampule doğru yukarı uçmadılar veya ışığın etrafında kümelenmediler, ancak gölgelik koridorun altından ışık altında çeşitli yollar izlediler (Şekil 4b), çarpma davranışının böceklerin yukarı yönü duygusu ile gerçek yerçekimi yönü arasındaki uyumsuzluğun bir sonucu olduğu fikrini destekliyor (Şekil 4c).
Daha küçük böceklerin sırt ışık tepkisinin manipülasyonuna karşı daha dirençli olup olmadığını test etmek için, çeşitli küçük böcekleri (vücut boyutu 2 cm veya daha az, 6 takımın 13 familyasından, taksonomik kompozisyon için Tablo 4'e bakın) yakaladık. Yüksek hızlı film çekimi için denekleri şeffaf bir Perspex küboid tankına (her tarafı 20 cm) yerleştirdik. Yukarıdan gelen difüz UV ışığıyla (~400 nm), test edilen tüm küçük böcekler normal kaçış uçuşuna benzeyen hızlı ancak istikrarlı bir şekilde muhafazanın tavanına doğru yukarı uçtu. Bununla birlikte, alttan gelen UV ışığıyla, Drosophila sp. hariç (daha sonraki bir bölümde ele alınacaktır) test edilen taksonlardan hiçbiri uçuşunu koruyamadı, kalkıştan kısa bir süre sonra eğildi ve ters döndü ve yere düştü (Şekil 4d, Ek Video 5). Bu sonuçlar, daha küçük böceklerin uçuşta yukarı yönünü belirlemek için ışığın yönüne büyük ölçüde güvendiğini ve dipteran halterleri gibi belirli duyusal organların dikeyliğin yanlış tahminini telafi etmediğini göstermektedir. Tüm Diptera ayrıca tanklarının üstüne ve altına soğuk beyaz LED ampullerle de test edildi. Hiçbir Diptera, beyaz kaynak üzerinde devrilme ve çarpma davranışı göstermedi, bu da etkinin test edilen Diptera'da kısa dalga boylu ışığa özgü olduğunu gösteriyor.
Normal uçuşun difüz bir gölgelik altında gerçekleştiği niteliksel gözlemlerimiz (Şekil 4b), 3B yeniden oluşturulmuş yörüngelerin bazı nicel ölçümleriyle doğrulanabilir (Ek Veri 5). Işığın yakınındaki yörüngeler için toplam yol kıvrımlılığı (toplam yol uzunluğu kat edilen mesafeye bölünerek), difüz bir gölgeliğin altından daha yüksektir (ortanca 3,21, n = 56) bir nokta kaynağının etrafında (ortanca 1,21, n = 56) (Wilcoxon rank sum Z = 6,32, p < 0,001) (Şekil 5a). Nokta ışık kaynaklarının yakınında uçan böcekler, ışığa dik olarak hareket etme eğilimindedir (Rayleigh z testi, z = 12,92, p < 0,001, n = 905), bu etki difüz gölgelik altında yoktur (z = 0,35, p = 0,70, n = 577) (Şekil 5b). Son olarak, ışık kaynağı böceğin hızının solunda veya sağındayken (zemin düzlemine yansıtıldığında dikliğin 30°'si içinde) ışık merkezli bir dönüş önyargısı olup olmadığını test ettik. Bir nokta ışık kaynağının yakınında kayıt edilen böcekler, beklendiği gibi ışığa doğru eğilmiş bir uçuş duruşunda olduğu için ışığa doğru tercih edilen bir dönüş sergilediler (X2 = 114,66, p < 0,001, n = 371) (Şekil 5c). Bu dönüş önyargısı difüz bir gölgelik altında mevcut değildi (X2 = 0,79, p = 0,37, n = 183).
Simüle edilmiş sırt eğimi, ışık tuzağına yakalanmayı üretmek için yeterlidir
Simülasyonda, alan ve laboratuvar ortamlarında gözlemlenen kalıpların yalnızca önerilen DLR mekanizmasından kaynaklanıp kaynaklanamayacağını test ettik. Çırpınarak uçuş için anatomik kısıtlamalar nedeniyle, uçan hayvanlar genellikle vücutlarına göre nispeten sabit bir oryantasyonda net bir aerodinamik ivme üretir37. Sonuç olarak, uçan hayvanlar genellikle yön değiştirmek için vücutlarını eğik tutar, yavaş uçuş manevraları (örneğin, havada durma) hariç33. Hareket yakalama verilerimizden aerodinamik ivmeyi (yerçekimini hesaba katarak) yeniden oluşturarak, net ivme vektörlerinin böceğin toraksına göre ileri ve sırtta dar bir aralıkta kümelendiğini bulduk (Ek Şekil 3).
Ajan tabanlı simülasyonlarımız, böceğin vücut eksenlerine göre sabit bir ivme vektörü kullandı (Şekil 6a). Uçuşun devam etmesi için toplam kaldırma kuvvetinin yerçekimine eşit veya ondan büyük olması ve ileri bileşenin seyahat hızı için sürtünmeyi önlemesi gerekir. Bu fenomenolojik modeli oluşturmak için doğrusal bir oransal denetleyici kullandık (ayrıntılar için Yöntemler bölümüne bakın). Dört serbest parametre vardı – k1: ışığa doğru sırt eğiminin kazancı, k2: gerçek dikeye doğru düzeltici sırt eğiminin kazancı, k3: vücut eksenini hız vektörüne doğru dengelemenin kazancı ve sürtünme büyüklüğünün bir indeksi olarak böceğin son hızı vt.
Üç davranışsal motifin her biri (Şekil 1), farklı başlangıç konumu verilen aynı parametre ayarlamalarıyla modelde çoğaltıldı. Uygun girişte, simüle edilmiş böcek, birkaç saniye boyunca sabit bir uçuş hızıyla ışığın etrafında yörünge uçuş yolu geliştirdi (Şekil 6b). Durgunluk, ajanın başlatılması ve ışık kaynağından uzağa uçması, dik bir ışıkla indüklenmiş tırmanma ve uçuş hızında azalma ile yeniden yaratıldı (Şekil 6c). Son olarak, ajanın girişi ışığın üstünden başlatıldığında, uçuşunu ters çevirdi ve hızla artan hızla bir dalışa girdi (Şekil 6d).
Üç motif sabit bir parametre ayarlamasıyla oluşturulurken, ışıkla indüklenen sırt eğiminin varsayımı, özel bir ayarlama yapılmadan ışık tuzağına kolayca neden olabilir. Rastgele parametreler ve başlangıç pozisyonlarıyla 300 beş saniyelik simülasyon çalıştırdık (Şekil 6e). Simüle edilmiş yörüngelerin çoğunluğu, ışık kaynağından korunmuş veya azalan menzil yoluyla ışık tuzağına yakalandığını gösterdi. Işıkla indüklenen sırt eğiminin kaldırılması (ışığı kapatmaya eşdeğer), ışık kaynağından dağılımı gösterdi (Şekil 6f). Her iki koşul altında da dağılımı, simülasyonun son 3 saniyesi için ışığa olan ortalama menzil değişimini ölçerek nicelleştirdik. Sırt eğim modellerinin ortanca menzil değişimi -0,06 m/s idi ve 0'dan istatistiksel olarak farklı değildi (Wilcoxon işaretli rank testi Z = 0,76, p = 0,45), bu da ajanların ışıktan kaçmadığını göstermektedir. Sırt eğimi olmayan modellerin ortanca menzil değişimi 1,85 m/s idi ve bu da ışıktan uzağa dağılımı göstermektedir (Wilcoxon işaretli rank testi Z = 14,42, p < 0,001). Deneylerde olduğu gibi, simüle edilmiş sırt eğimi modellerinin ortalama hız yönü ışığa dik olarak yer aldı ve dolambaçlı değil, dolaylı bir yol ile tuzağa düşmeyi vurguladı (Şekil 6g). Modelimiz, sırt eğiminin gözlemlediğimiz ışık tuzağına yakalanma uçuş yollarını oluşturmak için yeterli olduğunu göstermektedir.
Simülasyonumuzu, ışık tepkisi denetleyicisinin ışığı sabit, ancak keyfi bir öz merkezli konumda (sadece sırtta değil) tutması için değiştirdik. Bu model daha sonra yapay bir ışık kaynağının yakınlığı tarafından bozulmuş bir gök pusulasını temsil etti (Ek Şekil 4). 300 beş saniyelik rastgele parametre simülasyonunda, yörüngeler gerçek hayvanların gözlemlerimizle zayıf bir uyum gösterdi. Bazı hayvanlar ışık kaynağına doğru spiral halinde ilerlerken, yörüngeler hem gerçek böceklerde hem de DLR simülasyonlarında gözlemlenen tutarlı ışıkla dik yörüngelerden yoksundu. Gök pusulası simülasyonlarının ortanca menzil değişimi -1,75 m/s idi ve bu da ajanların ışıktan kaçtığını göstermektedir (Wilcoxon işaretli rank testi Z = 14,08, p < 0,001).
Işık değiştirme yoluyla uçuş yolu manipülasyonu
Bozulmuş bir pusula ipucu, böceklerin ışık kaynağının etrafında dairesel olarak (veya daha doğru bir şekilde logaritmik spiraller halinde) seyahat etmesine de neden olabilir8,19,38. Uçuş kontrol refleksi hipotezimizi klasik pusula navigasyon teorisinden kesin olarak ayırt etmek için, vahşi böcekler (tür kompozisyonu için Tablo 5'e bakın) ya her iki ışık kaynağının altında yörüngede dönerken iki farklı nokta UV ışık kaynağı arasında geçiş yaptık (Ek Şekil 5, Ek Video 6). 70 tek gözlü, yukarı bakan video topladık, bunlardan 37'si geçiş yaptıktan sonra yeni ışık kaynağının yörüngesinde dönen böcekleri içeriyordu. Diğer 33 kayıtta, böcekler ikinci ışık kaynağına yaklaşmadı. Göksel bir pusulanın karışıklığından tuzağa düşen böcekler, algılanan gök cismini aynı göreli konumda (sol veya sağ) tutmaya çalışırdı. Bununla birlikte, bir yönde (örneğin, saat yönünde) bir ışığın yörüngesinde dönen böceklerin, ışık kaynaklarını değiştirdiğimizde ışığa bakan taraflarını kolayca değiştirdiğini (ters saat yönüne geçtiğini) bulduk (böcekler 37 denemeden 25'inde yörünge yönünü değiştirdi). Ek olarak, 3 videoda, böcek ışık değiştirmeden aynı ışık kaynağında yörünge yönünü değiştirdi. Sırt eğimi, pusula navigasyonu için gerekli olan örtük S:S taraf sabitliğinden yoksun olan vücut yuvarlanması ayarıyla bu hızlı yön değiştirmeyi açıklar.
Işık tuzağına düşme davranışlarına istisnalar
Test edilen bazı böcek türleri ışık tuzağına yakalanmaya karşı bağışık gibi görünüyordu. Laboratuvar koşullarında, test edilen üç Zakkum Şahin Güvesi (Daphnis nerii)'nin hiçbiri 71 kayıtlı yörünge boyunca ışık yönlendirme davranışı göstermedi. Şahin güveleri, duruşlarını ters çevirmeden veya ışıkların yörüngesinde dönmeden yukarı bakan UV ve beyaz LED ampullerin hemen üstünden uçtular (Ek Video 7). Işığın yakınındaki D. nerii'nin yolları, diğer türlerde görülen dik eğilimden yoksundu (Ek Şekil 6). D. nerii için sırt eğim indeksi 0,24 (n = 911) idi ve karanlıktaki S. striolatum'unkinden zar zor büyüktü. Genel olarak, D. nerii, diğer türlerde bir nokta ışık kaynağının etrafında görülen aşırı eğim ve açılma açıları olmadan daha düz bir vücut duruşunu korudu (Ek Şekil 6). Vahşi yakalanmış sirke sinekleri (Drosophila spp.), bir UV veya beyaz LED ışık kaynağının üstünde veya altında uçuş arasında belirgin bir fark göstermeyen bir başka istisnaydı (Ek Şekil 6). Bu istisnalar, dalga boyu özgüllüğüne ek olarak, bu davranışta tür farklılıklarının da olduğunu düşündürmektedir. Bazı türler, hava duruşlarını yerçekimine göre düzeltmek için ışığa güçlü bir şekilde güvenmeyebilir.
Hayvan yetiştiriciliği
Laboratuvar deneylerinde kullandığımız böcekler ya alandan yakalandı (Sarı alt kanatlı güveler, Noctua pronuba ve Noctua fimbriata (sırasıyla n = 8, n = 2), Yaygın Okçu, Sympetrum striolatum, Göçmen Okçu, Aeshna mixta) ya da satın alınan pupalardan yetiştirildi (Atlas Güvesi Attacus lorquinii ve Zakkum Şahin Güvesi Daphnis nerii). Tüm böcekler, özel bir yetiştirme çadırında 24 °C ve %65 nemde 16–8 saatlik bir ışık-karanlık döngüsünde tutuldu. Yetişkin olarak beslenen güvelere (Saturniidae değil) yarı organik muz verildi. Yusufçuklar, laboratuvardaki birkaç gün boyunca elle beslenen yetişkin Drosophila spp. ile beslendi.
Yapay aydınlatma
Deneysel aydınlatmayı üç alternatif ampul türüyle sağladık. İlki, böcek ışık tuzaklarında yaygın olan mavi-UV tüp ışığıydı (Philips 15w TL-D Aktinik, Ek Şekil 7'ye bakın). İkincisi, bir UV LED ampul (TBE Lighting L276, 9w, Ek Şekil 7'ye bakın). Son olarak, soğuk beyaz bir LED ışık kaynağı kullandık (QNINE B22-G45 6000k, 6w). Bu ışıklar, böceklerde ışık tuzağına neden olan bir dizi ışık spektrumunu yansıtmak için seçildi. Bununla birlikte, bu çalışmada dalga boyunun etkilerini sistematik olarak takip etmedik. Işık kaynaklarımızın hiçbiri güçlü bir şekilde polarize değildi, bu da bazı böceklerde bulunan polarize ışığa doğru çekim etkilerini ortadan kaldırıyor51. Işık kaynakları laboratuvar deneylerinde çeşitli farklı konfigürasyonlarda kullanıldı. UV-Tüp ışıklar her zaman arenanın ortasına dikey olarak asıldı. UV ve Beyaz LED ampuller arenanın ortasına yerleştirildi ve ya yukarıya (ampul yukarı) ya da aşağıya (ampul aşağı) doğru yönlendirildi. Kontrol ışık ortamları oluşturmak için, gündüz türleri için geniş spektrumlu HID lambalar (2 x Philips CDM-TMW Elite 315 W çadır tavanından yayılmış) veya gece türleri için düşük yoğunluklu bir UV gölgelik oluşturmak için tavana parlayan tek bir gizlenmiş UV tüp kullandık.
Alan stereo videografi kayıtları
SINAC (Ulusal Koruma Alanları Sistemi) tarafından verilen M-P-SINAC-PNI-ACAT-024-2020 ve R-SINAC-ACG-PI-016-2022 numaralı izin belgeleri kapsamında, Estación Biológica Monteverde (EBM) ve CIEE, Monteverde Saha İstasyonu, Kosta Rika'da alan kayıtları yaptık (Ek Şekil 1, Ek Veri 1). Veriler iki ayrı saha gezisinde (Ocak-Şubat 2022 ve Mayıs 2023) toplandı. Tek bir üçayak çapraz koluna monte edilmiş bir çift monokrom deklanşör senkronize Fastec TS3 yüksek hızlı kamera kullandık. Çoğu video 500 fps hızında çekildi ve bu da uçuş davranışı için iyi bir zamansal çözünürlük sağladı. Bu kameralar, böcek gözlerine görünmez olduğunu varsaydığımız kızılötesi aydınlatma ile film çekmemize olanak sağladı. Bu varsayımla tutarlı bir şekilde, böceklerin kızılötesi ışıklarımıza (Larsen geniş açılı kızılötesi aydınlatıcılar; 850 nm) çarptığını veya bunlarla etkileşime girdiğini gözlemlemedik. Uçan böcekler ile karanlık gece arka planı arasında yüksek kontrast oluşturmak için kızılötesi aydınlatmayı yapılandırdık. Tam kamera yönleri ve ışık kaynağından olan mesafe geceye göre değişse de, her zaman ışık kaynağını her iki kameranın görüş alanının ortasına yerleştirdik. Uygulamada, bu bize maksimum 1,5 m x 2 m x 1,5 m'lik bir kayıt hacmi sağladı (Genişlik x Derinlik x Yükseklik, yükseklik yerçekimiyle hizalanmıştır). Stereo kalibrasyon sağlamak için, her iki kameranın üst üste binen görünümlerinden bilinen büyüklükte bir dama tahtası salladık. Daha sonra yerleşik MATLAB kamera kalibrasyon uygulamasını (Bilgisayar Görüşü Araç Kutusu 10.3) kullanarak hem her iki kameranın görünümlerindeki dama tahtalarını tespit etmeyi hem de hem içsel (optik merkez, odak uzaklığı ve radyal bozulma) hem de dışsal kamera parametrelerini (göreli kamera yönü ve öteleme) tahmin edebildik. Alan kayıtlarımızda, birçok böceği takım düzeyinin altından kesin olarak tanımlayamadık. Işıkların birkaç farklı konfigürasyonunu filme aldık. 'Işık Yok': Kameralar, UV veya görünür ampuller veya tüplerden herhangi bir aydınlatma olmadan ormanın bir uzay bölgesine yöneltildi. Kızılötesi aydınlatma hala sağlandı ve kırmızı renkte minimal sızıntı mevcuttu (Ek Şekil 7). 'UV Tüp': UV tüp ışığı, yerden yaklaşık 1,5 m uzaklıkta iki ağaç arasına ipten dikey olarak asıldı. Makro UV Tüp aynı işlemdir, ancak böcekleri gözlemlemek için verilerin analiz amacıyla her iki işlem de birleştirilmiş olan verilerin analiz amacıyla kameralar birkaç deneme için daha yakına taşındı. 'UV Ampul Yukarı': Yukarıda bahsedilen UV LED ampul, yerden yaklaşık 1 m yükseklikte üçayağa monte edildi ve yukarı doğru yönlendirildi. 'UV Ampul Aşağı': Aynı UV LED ampul, yerden yaklaşık 1,5 m yükseklikte asıldı. 'Beyaz Çarşaf Yukarı': Beyaz bir pamuk çarşaf yerden yaklaşık 2,5 m yükseklikte asıldı ve difüz olmayan aşağı doğru gelen ışığı önlemek için tabanında ek bir koruma kullanarak UV tüp ışığı ışığa yakın yukarı doğru yönlendirildi. 'Çarşaf Aşağı': Beyaz pamuk çarşaf orman zeminine serildi ve UV tüp ışığı yerden yaklaşık 0,3 m yükseklikte aşağıya doğru yönlendirildi. Işığa gelen farklı böceklerin daha iyi bir genel görünümünü elde etmek ve türlerin örneklemesini artırmak için, tek bir ışık koşuluyla (UV Ampul Aşağı) 6 takımda 30 böcek türünü içeren bilinen bir 30 böcek türü seti kullanarak 2023'te ek bir saha çalışmasıyla ışık çekim deneylerini tekrarladık. Böceklerin kimlikleri doğrulanmak üzere fotoğraflandı, ardından serbest bırakıldılar ve yukarıda açıklanan stereo kurulumla filme alındılar.
Gözlemlenen farklı motifleri (varlık-yokluk) ve videoda bulunan taksonu (farklı taksonomik seviyelere göre tanımlanmış) not ederek videoları niteliksel olarak inceledik. 'Yörünge': Hayvanlar ışığın etrafında yay şeklinde hareket ediyorsa (döngüler tamamlanmamış olsa bile), yörünge uçuş davranışı gösterdiği olarak sınıflandırıldı. Yeniden oluşturulmuş yörüngelerden, üstten bakıldığında dairesel veya oval yollardan döngüsel davranışın tanımlanması kolaydı. 'Durgunluk': Yukarı doğru uçan ve yukarı doğru eğilirken yavaşlayan ancak tamamen ters dönmeyen hayvanlar, durgunluk davranışı gösterdiği olarak sınıflandırıldı. 'Ters Dönme': Uçuşun herhangi bir bölümünde sırtlarını tamamen aşağı doğru eğen hayvanlar, ters dönme olarak sayıldı.
Birkaç videoda, uçuşun farklı bölümlerinde ve farklı türlerde birden fazla motif vardı, bu da nicelleştirmeyi zorlaştırdı ve bu nedenle videodaki tüm hayvanlarda görülen motifleri ekledik. Son sınıflandırma biraz özneldir, ancak gelecekte çapraz doğrulama için orijinal videoları son depoda sağlıyoruz. Bunu, motiflerin her izdeki yalnızca izlenen böcekler için tanımlandığı sayısallaştırılmış izler için de tekrarlıyoruz.
Laboratuvarda Hareket Yakalama İçin Böcek İşaretleme
Hareket yakalama sistemimiz, kayıt yapılan nesneye takılan retroreflektörlü işaretlere dayanmaktadır (Ek Şekil 2, Ek Veri 2). Retroreflektörlü işaret çerçevelerini (çerçeve başına 3 işaret) seri olarak üretmek için stereolitografik bir 3B yazıcı (Formlabs Form 3) kullandık. Daha sonra küçük (1 mm2) yapışkan retroreflektör bant (Qualisys) bölümlerini küresel işaretlere ekledik. Ortaya çıkan fotopolimer reçine işaret çerçeveleri, daha önce kullandığımız karbon çerçevelerden biraz daha ağırdı30 (işaret seti başına 10-20 mg), ancak üretim süresi çok daha kısaydı. Bu çalışmanın uçuş davranışı için, bu ağırlığın en hafif böceklerimiz için (%5 vücut ağırlığı) uçuş üzerinde minimal bir etkisi oldu (S. striolatum yaklaşık 300 mg). Denekler buza alınarak hareketsiz hale getirildikten sonra, minimum miktarda UV kürlenebilir yapıştırıcı (Loctite 4305) kullanarak toraksın sırt yüzeyine bir işaret çerçevesi taktık. Küçük bir ışık penceresi (3 mm) olan özel bir UV LED kalem, böceğin görüşünü hasar verme riskini en aza indirmek için yapıştırıcıyı yerel olarak kürlemek için kullanıldı. Kayıtlara başlamadan önce tüm böcekler davranışsal çadırda 20 dakika toparlandı. İşaretli ve işaretlenmemiş böcekler arasında gözle görülür davranışsal farklılıklar bulamadık, bu da işaret çerçevesinin genel uçuş kontrolünü etkilemediğini düşündürüyor.
Hareket yakalama davranış kayıtları
Çelik bir halkının (çap 1,66 m) (Ek Şekil 2) etrafına yerleştirilmiş kızılötesi (850 nm) kayıt yapan sekiz Qualisys Marqus M5 hareket yakalama kamerası (4 çift) kullandık. Bu, tavandan asılı dikey olarak hareket ettirilebilir bir metal çerçeveye (2,4 × 2,4 m) tutturuldu ve hem yükseltildi hem de merkezi bir vinç tarafından alçaltıldı. Sonuçlarımızı etkileyen laboratuvardaki ışık sızıntılarını (örneğin, bilgisayar ekranları) önlemek için karartma perdeleri kullandık. Bu çerçeveden beyaz silindirik bir çadır (çap 2 m, yükseklik 2,4 m) asılıydı ve kameralar tavana yakın yerleştirilmiş pencerelerden dışarı bak