Bugün öğrendim ki: Neandertallerin soğuk hava koşullarında hayatta kalmak için bazı modern insan topluluklarına ısı üretmek için kas kütlesi, tıknaz yapı, büyük burun ve sinüsler gibi genetik özellikler aktardığı ve bu özelliklerin doğal olarak seçildiği anlaşılıyor.
Tropikal ve sıcak çöl bölgelerinden daha düşük sıcaklıklara uyum sağlama, anatomik olarak modern insanların Afrika dışı göçlerinden sonra Avrasya kara kütlesinin kolonizasyon süreçleri sırasında son 100.000 yılda insan türünde makul bir şekilde geliştirilmiştir. Bazı insan gruplarının orta ve boreal ortamlarda binlerce yıllık yerleşimi, morfolojik ve kültürel özelliklerini şekillendirmiştir, ancak aynı zamanda metabolik adaptasyonların, soğuktan ve vücut ısı üretiminin izolasyonunu desteklediği varsayılmaktadır (Leonard ve ark., 2002).
Bu görüntü, Neandertaller ve belki de bazı Denisovan popülasyonlarına da genişletilebilir, çünkü hem paleoantröpolojik kalıntılar hem de genetik veriler, görünüşlerinin ve yok oluşlarının çoğunlukla son buzul çağının şiddetli bir şekilde yaşandığı coğrafi alanlarda, çoğunlukla son buzul döneminde gerçekleştiğini (Lalueza-Fox ve Gilbert, 2011) ve bunun için düşük ortalama sıcaklıkların tahmin edildiğini (Skrzypek ve ark., 2011) göstermektedir. Buna göre, Neandertal popülasyonlarının çoğunun ve muhtemelen bazı Denisovan popülasyonlarının da atalarının yaşadığı sıcaklıklara göre önemli ölçüde daha düşük sıcaklıklarla başa çıktığı ve bunlara uyum sağlayacak kadar uzun bir zaman diliminde kaldığı varsayılabilir. Neandertaller özellikle, muhtemelen termal verimliliğini artıran (Holliday, 1997) veya soğuktan kaynaklanan damar daralması nedeniyle daha kötü bir kan dolaşımının bir sonucu olarak yorumlanabilen morfolojik özellikler sergilemişlerdir (Steegmann, 2007). Ayrıca, varsayılan büyük kas aparatı ve avcı-toplayıcı geçim stratejileri, yaşam tarzlarının kutup bölgelerindeki modern popülasyonlarınkine benzer olabileceğini, en azından yüksek enlemlerde yağ ve et proteinleri bakımından zengin bir diyetle karakterize olduğunu (El Zaatari ve ark., 2011) ve yüksek kalori ihtiyacını karşılayabileceğini (yani, günde 4000-7000 kalori) (Snodgrass ve William, 2009) göstermektedir. Bu özellikler, modern popülasyonlarda düşük sıcaklıkla genellikle ilişkilendirilen yüksek bazal metabolik hızla (Leonard ve ark., 2002) eşleştirilerek bu antik türde soğuk uyumunu geliştirmiş olabilir.
Özellikle, tropikal olmayan ortamlara metabolik adaptasyonlar, hipotalamusun soğuk maruziyete yanıt olarak ürettiği sempatik sinyaller tarafından kontrol edilen karmaşık fonksiyonel yollar olan termojenez ve termoregülasyon süreçlerinin modülasyonu tarafından yönlendirilebilir. Bu sinyaller, trigliseritlerden serbest yağ asitlerinin salınımını tetikler, termojenini aktive eder, oksidatif fosforilasyonu ve ATP sentezini koparır ve mitokondride üretilen enerjinin ısı olarak dağılmasına yol açar (Wijers ve ark., 2008). Kahverengi yağ dokusu (KJD) adipositeleri, beyaz yağ adipositelerine kıyasla daha fazla mitokondri miktarı ve iç zarında daha yüksek termojenin konsantrasyonuna göre bu titreme olmayan termojenezin en özelleşmiş hücreleridir (Richard ve Picard, 2011). Aslında, KJD, özellikle kış uykusuna yatan memelilerde bol miktarda bulunur, ancak esas olarak insan türünün ötesinde en geniş coğrafi dağılıma sahip primatlardan biri olan Macaca mulatta'da bazı maymunlarda da bulunmuştur. Dahası, birkaç yıl öncesine kadar Homo sapiens'te KJD varlığının kanıtı sadece yenidoğanlarda sınırlı iken, son zamanlarda yetişkinlerde de gözlemlenmiştir (Cypess ve ark., 2009; Virtanen ve ark., 2009) ve soğuk maruziyete yanıt olarak daha metabolik olarak aktif görünmektedir (Saito ve ark., 2009; van Marken Lichtenbelt ve ark., 2009).
Dolayısıyla, KJD metabolizması, depolanması ve neojeneziyle ilgili genler, insan soyunda da termojenez ve termoregülasyon için sorumlu ana yollardan biridir. Bu genlerdeki genetik varyasyon, hem modern hem de antik insanlarda düşük sıcaklıklara metabolik adaptasyonlar ve gelişmiş bir termal verimlilik için önemli anahtarları temsil etme potansiyeline sahiptir.
Bu hipotezi araştırmayı amaçlayan keşifçi bir analiz yapmak için, farklı iklim bölgelerinden popülasyonlarda, yanı sıra Neandertal ve Denisovan genomlarında olası işlevsel etkiye sahip varyasyon modellerinin, insan vücut ısı üretiminde ve atalarımızdakilerde yer alan mekanizmaların evrimine yeni bilgiler katmaya yardımcı olmasını umarak araştırılmıştır.
Araştırılan genom bölgeleri
KJD metabolizmasında doğrudan rol alan veya üzerine olası düzenleyici işlevleri olan 28 gen panelinden (Çizelge 1), hassas literatür incelemesi ve String veritabanında (http://string-db.org/) bildirilen protein-protein etkileşimlerini keşfederek seçildi. Panel, hala bilinmeyen biyolojik işlevlere sahip, ancak geniş çaplı ilişki çalışmaları sonuçlarına göre yağ birikimi ve vücut kitle endeksi ile ilişkili genleri de içerir (www.genome.gov/gwastudies).
Kullanılan veri setleri
50x ve 30x kapsamlı Neandertal ve Denisovan genomlarından hg19 insan referans dizisine hizalanan dizi verileri sırasıyla http://cdna.eva.mpg.de/neandertal/altai/ ve https://bioinf.eva.mpg.de/download/HighCoverageDenisovaGenome/DenHC_catalog/ adreslerinden alındı. Seçilen genlerin eksonlarını kapsayan okumalar, özel olarak geliştirilmiş Phyton betikleri kullanılarak çıkarıldı ve nükleotid ikamelerini aramak için kullanıldı.
Modern popülasyonlar için, dikkate alınan genom bölgelerinde bulunan tek nükleotid polimorfizmleri (SNP'ler), 1000 Genom Projesi 1. faz veri kümesinde yer alan 14 insan grubuna ait 1092 bireyde tespit edilenlerden alındı ve yüksek kapsamlı ekzom hizalamalarının incelenmesiyle de doğrulandı (ftp://ftp-trace.ncbi.nih.gov/1000genomes/ftp/phase1/).
SNP tanımlama
SAMtools paketi (Li ve ark., 2009) nin bcftools yardımcı araçları, 1000 Genom Projesi SNP'lerini tanımlamak için kullanılan aynı parametre ayarlarını uygulayarak toplanan eski genom okumalarından varyantları çağırmak için kullanıldı (1000 Genom Projesi Konsorsiyum et al., 2012), mükemmel şekilde karşılaştırılabilir modern ve eski veri setleri elde etmek amacıyla. Neandertal ve Denisovan genomlarında sırasıyla tespit edilen varyantlar, modern veri kümesinde arandı ve ayrıca modern SNP'lerle ilgili genom konumları, eski genomlarda çapraz kontrol edildi. Buna göre, insan türünde dbSNP Yapı 137'ye göre zaten kaydedilmiş ve atalarının allelleri şempanze genomuyla karşılaştırılarak çıkarılmış 406 SNP'den oluşan bir veri kümesi elde edildi.
Modern varyasyondaki özet istatistikler ve popülasyon yapısı analizleri
SNP'lerin tamamı, her mevcut insan grubu için Arlequin paketi v.3.5.1.2 kullanılarak özet istatistikler (nükleotid çeşitliliği (π), lokuslar arasında ortalama beklenen heterozigotluk (MEH) ve polimorfik site sayısı (S)) hesaplamak için kullanıldı.
Ayrıca, bu veri kümesi, antik varyasyonu modern varyasyon içine yerleştirmeyi amaçlayan çeşitli popülasyon yapısı analizleri (Ek Materyaller ve Yöntemler) için kullanıldı. Bu amaçla, veri kümesi, çok değişkenli ve karışım analizlerinin yanı sıra kullanılan model tabanlı kümeleme yönteminin LD etkilerini önlemek için PLINK v.1.07 (Purcell ve ark., 2007) kullanılarak filtrelendi ve tek örnekler çıkarıldı ve SNP'ler bağlantı dengesine (LD) göre budandı. Bağlantı dengesi, 50 SNP'den oluşan kayan pencereler kullanılarak hesaplandı ve her iki SNP çifti arasında bağlantı dengesi hesaplandı; eğer iki SNP'nin genotipi korelasyonu (r2) > 0.1 ise, bu SNP çiftlerinden biri çıkarıldı. Her pencere daha sonra 10 SNP ileriye kaydırıldı ve aynı prosedür tekrarlandı, bu sayede birbirleriyle yaklaşık bağlantı dengesinde olan 244 SNP'den oluşan bir alt küme elde edildi.
Popülasyon gruplarının farklılaştırılması analizleri
Yapı analizleriyle belirlenen genetik olarak homojen popülasyon grupları arasındaki farklılaşma modelleri (Ek Materyaller ve Yöntemler), örnekleri baskın köken bileşenlerine göre gruplandırarak araştırıldı, örneğin Afrika (AFR), Avrupa (EUR), Doğu Asya (EAS) ve Latin Amerika (AMR). Araştırılan 406 SNP'nin her birinde, karşılaştırılan grupların her ikisinde de tek morfik olanlar çıkarıldıktan sonra, her bir 406 tanımlanmış SNP için Fst indeksini hesaplayarak araştırılan genler arasında en farklı olan genlerin tespiti gerçekleştirildi. Ayrıca, örnekler arasındaki alel frekanslarını karşılaştırmak için PLINK v.1.07 kullanılarak bir χ2 testi yapıldı ve χ2 ve Fst değerleri arasında gözlenen yüksek korelasyona göre, her bir SNP'nin farklılaşma düzeyiyle ilgili istatistiksel anlamlılık kanıtı sağlamak amacıyla kullanıldı. Elde edilen asimptotik p-değerleri için, kullanılan çoklu test prosedürlerine uyum sağlamak amacıyla, R paketi multtest kullanılarak Bonferroni düzeltmesi için düzeltilmiş p-değerleri hesaplandı.
Seçim için en olası hedefleri belirlemek için, SNP'ler küresel küçük alel frekanslarına göre bölümlere ayrıldı ve her bölgede Fst değerlerine göre sıralandı. Ardından, her bölgedeki Fst dağılımının üst %1'inde puan alan varyantlar, sonraki analizler için tutuldu. Bu yaklaşım, aynı frekans aralığına ait olanlar arasında potansiyel dışsılar olan, orta Fst değerine sahip SNP'leri de hesaba katmayı mümkün kıldı.
Belirlenen aday SNP'lerin ampirik genom çapı Fst dağılımları içerisindeki konumları, genomiğin örüntülerine kıyasla alışılmadık derecede yüksek farklılaşmaya sahip genleri kesin olarak belirlemek için araştırıldı. Özellikle, 1000 Genom Projesi popülasyonlarının dört kıtasal grubunu karşılaştırmak için özel olarak geliştirilmiş bir Perl betiği kullanılarak altı Fst genom çapı dağılımı elde edildi ve 99. yüzdeliklerinde puan alan SNP'ler anlamlı dışsılar olarak kabul edildi.
Aday lokuslar üzerindeki nötrlük testleri
Farklılaştırma analizinden elde edilen en iyi aday lokuslar için, HapMap Faz II ve HGDP veri kümelerinde hesaplanan Entegrasyon Haplotip Skoru (iHS) değerleri, Haplotter (http://haplotter.uchicago.edu/) ve HGDP Seçim Tarayıcısı (http://hgdp.uchicago.edu/cgi-bin/gbrowse/HGDP/) web uygulamaları kullanılarak alındı. Bu lokuslar için tam gen dizisi verileri, 1000 Genom Projesi 1. faz veri kümesinden de alındı ve çeşitli özet ve nötrlük istatistiklerini hesaplamak için kullanıldı. Özellikle, DnaSP paketi v.5.10, ortalama ikili fark sayısını (Pi), Watterson'ın nükleotid çeşitliliği tahminleyicisini (θW), Tajima'nın D, Fu ve Li'nin D* ve F* ve Fay ve Wu'nun H'sini hesaplamak için kullanıldı. 10 kb'lik örtüşmeyen kayan pencerelerde. Yerel rekombinasyon oranı ve Gravel ve ark. (2011) tarafından tanımlanan demografik modeller üzerinde koşullandırılan 10.000 koalesan simülasyonu yapmak için Fastsimcoal paketi v.2 (Excoffier ve ark., 2013) kullanıldı. Yukarıdaki istatistikler daha sonra DnaSP paketi v.5.10 kullanılarak simüle edilmiş veri kümelerinde hesaplandı ve evrimsel bir nötr model altında değerlerin dağılımlarını üretmek için kullanıldı. Her istatistik için tahminler, elde edilen dağılımlarla tek yönlü testlerde karşılaştırıldı ve anlamlılık, gözlemlenenlerden daha aşırı değerlere sahip koalesan simülasyonlarının oranı olarak alındı. Uygulanan p-değerleri için, kabul edilen çoklu test prosedürlerini kontrol etmek amacıyla, R paketi multtest kullanılarak uyarlanmış Benjamini ve Hochberg (ABH) prosedürü (Benjamini ve ark., 2006) için düzeltilmiş p-değerleri hesaplandı.
LD, haplotip ve ağ analizleri
1000 Genom Projesi 1. faz veri kümesinden, en iyi aday genin tamamını, yanı sıra 100 kb yukarı ve aşağı bölgeleri kapsayan bir genom aralığı için dizi verileri alındı ve her bir olası SNP çifti için ikili LD hesaplamak için PLINK v.1.07 kullanıldı.
En umut verici aday varyantla yüksek LD (r2> 0.95) içindeki SNP'ler, PHASE v.2.1'de uygulanan Bayesçi algoritma kullanılarak haplotiplerin yeniden yapılandırılması için kullanıldı. Çıkarılan haplotiplerin evrimsel ilişkileri, Network paketi v.4.6.1.1 (http://www.fluxusengineering.com) kullanılarak orta katılımlı bir ağ kullanılarak görselleştirildi.
Bu çalışmada kullanılan betikler, ilgili yazara başvurarak temin edilebilir.
KJD genlerinin dizi çeşitliliği
Modern veri kümesinde, dbSNP Yapı 137'ye göre kaydedilmiş toplam 406 SNP gözlendi (Ek Tablo 1), bunlardan 12'sinin türetilmiş allellerinin antik türlerde de bulunduğu görüldü. Özellikle, hem Neandertal hem de Denisovan genomlarında, tüm modern popülasyonlarda yüksek ve orta frekansta (sırasıyla ≥76%, ≥84%, en fazla %34%) görülen PLIN1 rs6496589 ve PPARGC1B rs7732671'in yanı sıra PLIN1 rs2304796 allelleri gözlendi; sadece birkaç AFR bireyinde (1%) bulunan PLIN1 rs150128694 ve EAS'te (87%) yüksek oranda temsil edilen LEPR rs1137101. Diğer altı SNP'nin türetilmiş allelleri yalnızca Neandertal genomunda gözlendi; LEPR rs6413506 sadece birkaç AFR bireyinde (%4) bulundu; PPARG rs3856806 ve PPARGC1B rs45520937, rs45588534 ve rs45543631 dünya çapında orta frekansta (en fazla %20) bulundu ve NRF1 rs3735006 tüm modern popülasyonlarda düşük frekansta (en fazla %7) gözlendi. Son olarak, sadece birkaç AFR bireyinde (%2) bulunan PPARG rs41516544 türetilmiş aleli, Denisovan genomunda heterozigot durumda bulundu.
Modern varyasyondaki özet istatistikler ve popülasyon yapısı
Modern popülasyonlar için dizi çeşitliliğinin özet istatistikleri Ek Tablo 2'de sunulmaktadır ve her popülasyonun kısaltmaları, Ek Şekil 1'in efsanesinde de tekrarlanmıştır. Beklendiği gibi, baskın AFR kökenli etnik grupların çeşitliliği, karışım göstermeyen Avrasya popülasyonlarına kıyasla daha yüksekti ve bu da polimorfik lokusların önemli ölçüde daha yüksek yüzdesine ilişkilendirildi. Bununla birlikte, en yüksek heterozigotluk değerleri, İspanya'dan İber popülasyonlarında (IBS) alışılmadık bir tepe hariç, EAS ve kuzey EUR popülasyonlarında bulundu; bu, İber Yarımadası'ndan farklı popülasyonlardan bireylerin dahil edilmesi ve küçük örneklem büyüklüğü nedeniyle yüksek heterojenlik ile açıklanabilir.
Çift popülasyon karşılaştırmalarının büyük çoğunluğu için önemli Fst genetik uzaklıkları hesaplandı, birkaç istisna hariç (Pekinli Han Çinlileri, CHB karşı Güney Çinli Han Çinlileri, CHS, P=0,714; Medellínli Kolombiyalılar, CLM karşı Porto Rikoluklar, PUR, P=0,188; Kuzey ve Batı Avrupa kökenli Utah sakinleri, CEU karşı İngiltere ve İskoçya'dan İngilizler, GBR, P=0,238 ve IBS'yi içeren çoğu karşılaştırma). Elde edilen ikili uzaklık matrisi (Ek Tablo 3), gözlenen varyasyon için önemli bir coğrafi yapıyı ortaya koyan ve gözlenen popülasyon varyasyonunun yaklaşık olarak kıtasal gruplara karşılık gelen dört kümenin varlığını gösteren bir metrik çok boyutlu ölçeklendirme (MDS) ile grafiksel olarak temsil edildi (Ek Şekil 2). Tekrar, IBS örneği, diğer EUR'lere göre CLM ve PUR'a daha yakın görünüyordu. Bu desen, dünya çapındaki popülasyonlar arasında anlamlı ancak nispeten düşük bir farklılaşma gösteren (FST=0,075, P<0,001) ve örnekler coğrafi temelli kümelere ayrıldığında, MDS grafiğinin önerdiği gibi IBS'nin AMR popülasyonlarıyla kümelenmesi durumunda bile azalan orta ve anlamlı gruplararası varyans bileşeni (FCT=0,081, P<0,001) gösteren moleküler varyans (AMOVA) sonuçları (Ek Tablo 4) ile istatistiksel olarak desteklendi (FCT=0,079, P<0,001).
Eski varyasyonu modern varyasyon bağlamına yerleştirme
Eski profilleri modern varyasyon bağlamına yerleştirmek için, Neandertal ve Denisovan verilerini de içeren ve birbirleriyle yaklaşık bağlantı dengesinde olan 244 SNP alt kümesini kullanan daha fazla popülasyon yapısı analizi yapıldı.
Karışım analizi için en iyi tahmin gücüne sahip model, baskın AFR, EUR ve EAS kökenli popülasyon kümelerine yaklaşık olarak karşılık gelen üç atasal grup önerdi; EUR ve AMR popülasyonları aynı kümeye atandı (Ek Şekil 1). K=3'te antik genler baskın bir AFR benzeri köken bileşeni (%62,8) ve daha sınırlı bir EAS benzeri bileşen (%37,2) gösterdi. Bu işaretler hem düşük hem de yüksek çözünürlükte gözlemlenebilirdi (sırasıyla K=2'de %23,9 ve K=4'te %38,7) ve K=5'te ortadan kayboldu (Ek Şekil 3).
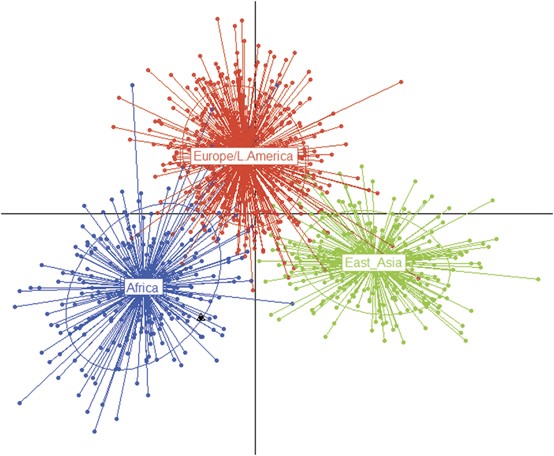
Modern veri ve antik örnekler için hesaplanan 20 ana bileşenden (PB) modern veri için uygulanan ayrımcı ana bileşen analizi (DAPC), antik örnekler "ek örnekler" olarak temsil edildi. AFR kümesi en düşük ortalama üyelik olasılığını (%0,898) gösterdi, en küçük tek değerleri arasında Güneybatı Amerika Birleşik Devletleri'nden (ASW) gelen Afrika kökenli kişiler vardı, bunların %29,5'i EUR/AMR kümesine atandı, EAS ve EUR/AMR kümeleri için değerler dikkate değer ölçüde daha yüksekti (%0,951 ve %0,966).
Antik örnekler, esasen AFR kümesinin parçası olarak tahmin edildi (%0,989 AFR üyelik olasılığı) ve EAS ve EUR/AMR bileşenleri için sırasıyla %0,001 ve %0,009 olasılıklar gösterdi (Şekil 1).
Son olarak, model tabanlı bir kümeleme analizi, yaklaşık olarak "AFR" (C1), "EUR/AMR" (C2) ve "EAS" (C3) kümeleri olarak tanımlanabilen üç kümenin varlığını doğruladı (Ek Şekil 4). Özellikle, incelenen AFR bireylerinin %94'ü, EUR'lerin %5,3'ü, AMR'lerin %7,2'si ve EAS'lerin %0,7'si C1 kümesini oluşturdu. C2, EUR örneklerinin %67'sini ve AMR örneklerinin %63'ünü, yanı sıra sırasıyla AFR'lerin %4,1'ini ve EAS'lerin %1,1'ini içeriyordu. C3, incelenen EAS örneklerinin %98'ini, AMR'lerin %30'unu, EUR'lerin %28'ini ve AFR'lerin %2'sini içeriyordu.
Modern popülasyon gruplarının farklılaştırılması
Yapı analizleriyle belirlenen genetik olarak homojen popülasyon grupları arasındaki en farklı genlerin tespiti, her bir SNP için Fst indeksini ve yüksek korelasyonlu χ2 testini (örneğin, AFR-EAS karşılaştırması için r=0,992, P<0,0001) hesaplayarak elde edildi. Ardından, her frekans bölgesindeki üst %1 Fst kuyruğu ve hesaplanan Fst genom çapı dağılımlarının %99. yüzdeliklerinde puan alan varyantlar, olağandışı derecede farklılaşmış lokuslar olarak işaretlendi.
Afrika dışı göçten sonra türetilmiş allellerinin yüksek frekansına göre uyarlanabilir bir soğukla ilgili rol oynamış olabilecek dışsal SNP'ler yalnızca EAS'yi içeren karşılaştırmalarda tanımlandı. Aslında, AFR-AMR ve EUR-AMR karşılaştırmalarında yüksek derecede farklılaşmış alleller, Afrikalılar dışındakilerde yalnızca düşük ila orta frekanslar gösterdi (AMR'de en fazla %0,392 ve EAS'te en fazla %0,339 PRKAR2B rs75385144 ve FTO rs2287142 için) (Çizelge 2).
Buna göre, LEPR, kalan gruplardan ve sırasıyla AMR ve EUR'den EAS'i önemli ölçüde ayıran üç varyant (rs1137100, rs1137101 ve rs1805096) göstererek en umut verici aday gen olarak ortaya çıktı (Çizelge 2). Özellikle, rs1137100 türetilmiş aleli AFR'de çok düşük frekansta (%0,128), EUR'de ve AMR'de orta frekansta (%0,289 ve %0,265) ve EAS'te yüksek frekansta (%0,811) gözlendi. rs1137101 türetilmiş aleli de antik genomlarda gözlendi ve EAS'te daha da yüksek bir frekansta (%0,867) görüldü ancak kalan popülasyonlarda da önemli ölçüde temsil edildi (Çizelge 2). Aksine, rs1805096 türetilmiş aleli, muhtemelen rs1137101 atasal aleli ile önemli bir LD (r2=0,76) nedeniyle, EAS hariç tüm incelenen gruplarda yaygındı.
LEPR nötrlük testleri ve haplotip analizleri
LEPR geni üzerindeki olası doğal seçim işaretlerini daha fazla araştırmak için, ilgili 220.858 bp'lik genom bölgesini kapsayan 10 kb'lik örtüşmeyen pencerelerde nükleotid çeşitliliğinin modelleri değerlendirildi.
Her popülasyon grubu için elde edilen düşük ila orta Pi ve θW değerlerine (ayrık sitelerin düşük sayısıyla paralel olarak) rağmen, üç aday LEPR varyantını içeren 10 kb'lik genom pencereleri için koalesan simülasyonları kullanılarak hesaplanan p-değerlerine göre olağanüstü düşük nükleotid çeşitlilikleri gözlemlenmedi (Çizelge 3).
Yanlış keşif oranını kontrol ettikten sonra, rs1137100'ü kapsayan pencere söz konusu olduğunda, EAS için anlamlı ve büyük ölçüde negatif Fu ve Li'nin D* ve F* değerleri elde edildi (−5.056, P=0.0007, düzeltilmiş P=0.041 ve −3.049, P=0.002, düzeltilmiş P=0.044); bu, bu SNP çevresindeki alel frekans dağılımının nötrlük altında beklenen kalıplardan önemli bir sapmayı gösteriyor. Özellikle, Fu ve Li'nin D* ve F* tahminlerinin ancak Tajima'nın D'nin değil anlamlılığı, bu gruptaki tek örneklerin, diğer gruplardan ziyade yalnızca düşük frekanslı varyantlar yerine aşırı oranda temsil edildiğini gösterdi. Aslında, rs1137100 penceresinde, toplam ayrık sitelerin %41'ini oluşturan bu grupta gerçekten diğer gruplara göre daha fazla temsil ediliyordu.
Daha sonra LEPR genine göre 100 kb yukarı ve aşağı uzanan bir genom bölgesinde LD kalıpları incelendi ve bununla yüksek LD (r2≥0,95) içindeki 10 intronik SNP'nin tanımlanmasına yol açtı. Bu varyantlar, modern popülasyonları karakterize eden LEPR haplotiplerini yeniden yapılandırmak ve evrimsel ilişkilerini orta katılımlı bir ağ kullanılarak görselleştirmek için kullanıldı (Şekil 2). Toplam 20 çıkarılan haplotipten sadece ikisi (%96 örnek kromozom) hesaba katıyordu; kalanlarının uyarlanabilir bir işlevi olma olasılığı düşük. Bu yaygın haplotiplerden H20, her SNP için atasal aleli içeriyordu ve tüm kıtasal gruplarda (frekansı %58'i aşan) ancak EAS'te baskın haplotipti. Aksine, H1 her SNP için türetilmiş aleli taşıyordu ve tüm incelenen gruplarda orta frekansta (%36'ya kadar) gözlendi, ancak EAS hariç, burada kromozomların yaklaşık %60'ını oluşturdu ve potansiyel olarak uyarlanabilir bir rol oynadı.
Yukarıda belirtildiği gibi, kalan haplotipler tüm kıtasal gruplarda %1-2'den düşük frekans göstererek son derece nadirdi. Bunlardan altı tanesi (H2, H3, H6, H12, H14, H16), rs1137100 türetilmiş alelini taşıyordu ancak gözlenen topolojinin farklı dallarında yer alıyordu. Özellikle, H3, H2, H1 ve H6'nın tek, iki ve üç adım türevleri olarak ortaya çıktığı haplotip gibi görünüyordu, oysa H12, H14 ve H16 atasal haplotip H20'ye (Şekil 2) daha yakın görünüyordu. Buna göre, rs1137100'e ek olarak, gözlenen topolojide tekrarlanan diğer varyantlar ortaya çıktı; bu muhtemelen rekombinasyon veya gen dönüşümleri nedeniyle gerçek homoplasilerden ziyade oldu.
Dolayısıyla, çeşitli ek yaklaşımlar, LEPR genindeki varyasyonun, incelenen diğerler arasında, potansiyel olarak EAS popülasyonlarının tropik olmayan iklimlere metabolik adaptasyonuna gerçekçi bir katkıda bulunabilecek tek varyasyon olduğu konusunda kanıtladı. Bu gen, hipotalamik bölgelerdeki leptin bağlanması ve noradrenalin salınımı için sorumlu reseptörü kodlamaktadır; hedefleri, yağ dokusundaki yağ asidi işlenmesiyle ilgili sinyal yollarını başlatan ADRB3 ve ADRA1A genleridir. Buna göre, beslenme davranışını, yağ depolama ve glikoz metabolizmasını düzenleyerek tokluk ve enerji dengesini düzenler, aynı zamanda leptinin insülin salgılanmasını inhibe etmesine izin verir (Myers, 2004). Ayrıca, özellikle sempatik sinir aktivitesini artırarak KJD'deki norepinefrin dönüşümünü de artırır (Haynes ve ark., 1997).
Özellikle, EAS'te seçilim göstergesi olan LEPR SNP'lerinin bazı spesifik metabolik desenler ve/veya bozukluklarla ilişkili olduğu bildirildi. Örneğin, rs1137100, solunum oranı (yani, bazal metabolik hız) artışı ile ilişkili bulunan (K109R) bir anlamlı olmayan ikame için sorumludur (Loos ve ark., 2006), bu da titreme olmayan termojenezin önemli etkisine uygundur. Türetilmiş alelinin ayrıca glikoz toleransını ve insülin yanıtını etkilediği, bozuk glikoz toleransı olan kişilerde açlık insülin seviyeleri ve insülin yanıtları ile ilişkili olduğu bildirilmiştir, bu da tip II diyabet patojenisinde rol oynayabileceğini gösterir (Wauters ve ark., 2001). Aslında, insülini leptin tarafından inhibe etmeyi azaltır, insülin seviyelerinin düzenlenmesinde başarısızlığa ve artan insülin salımına yol açar, bu da glikoz alımını ve bazal metabolik hızı hızlandırır (yani, ısı dağılımını destekler), ancak potansiyel olarak insülin direnci durumuna da yol açar. Buna göre, bu varyantın bazal metabolik hızda ve dolayısıyla titreme olmayan termojenezin temeli olan ısı dağılımında artışa neden olduğu soğuk iklimlerde olumsuz bir etkisi vardır ve bu nedenle soğuk iklimlerde avantajlı hale gelir.
Modern insanlarda rs1137101 üzerindeki olumlu seçilim eylemi, genotipleme verilerine göre ancak dizi verilerinden elde edilen sonuçlarla sorgulanabilir (Fu ve Li'nin D*=−4,5134, düzeltilmiş P=0,068 ve Fu ve Li'nin F*=−2,379, düzeltilmiş P=0,069) varsayılabilir. Sonuç olarak, türetilmiş alelinin antik genomlarda da görülen ve kanıtlanmış işlevsel bir etkiye sahip tek modern alel olması, bu SNP'yi çok ilginç kılar. Özellikle, artan insülin salımı, vücut ağırlığı, vücut kitle indeksi ve özellikle Pasifik Adaları'ndaki birçok popülasyon için obezite riski ile ilişkili olduğu bulunan (Q223R) bir anlamlı olmayan ikame için sorumludur ve etkisi diğer genetik ve/veya çevresel faktörlere büyük ölçüde bağlı olduğu varsayılmaktadır (Furusawa ve ark., 2010). Modern ve antik genomlar arasında paylaşılan tek potansiyel soğuk uyarlanabilir aleli olmasına rağmen, tek Denisovan ve Neandertal bireylerin incelenmesi bu türlerin soğuk uyumuna ilişkin güvenilir sonuçlar çıkarmayı engelliyor. Özellikle, incelenen örneklerin geldiği (örneğin, güneydekilere göre Sibirya'daki gibi) soğuk bölgelerdeki antik popülasyonlarda bu SNP'nin gerçekten aşırı oranda temsil edilip edilmediğini test etme imkanımız yok. Aslında, modern AFR'de önemli bir frekansta (%0,539) bulunup her iki antik türde de var olan rs1137101, insan soyunda eski zamanlarda ortaya çıkmış olabilir ve bu, muhtemel rolüne ve Denisovan'ların varsayılan alt-tropikal Güney Doğu Asya kökenine bakılmaksızın tek şimdiye kadar sıralanan Denisovan örneğinde bulunmasını açıklayabilir (Meyer ve ark., 2012).
Dolayısıyla, bazı Neandertal ve Denisovan popülasyonlarının modern insanlarda yer alanlardan farklı işlevsel yollar aracılığıyla soğuk ortamlara uyum sağlamış olma olasılığı en olası görünmektedir.
Bu bağlamda, insan memelilerinde yapılan transkriptomik analizlerle ortaya çıkan yeni bulgularla uyumlu olarak, seçim etkisini titreme termojenesiyle ilgili yollarda da ayırt etme, düşük sıcaklıklara insan uyumunu araştırmak için tamamlayıcı ve umut verici bir yaklaşım olabilir. Aslında, Cheviron ve ark. (2012), yüksek irtifa geyik farelerinde, yağ asidi oksidasyonunda ve oksidatif fosforilasyonda yer alan genlerin önemli değişiklikleri aracılığıyla lipitleri temel metabolik yakıt kaynağı olarak kullanma kapasitesinin artmasının seçildiğini öne sürdü. Bu mekanizmanın titreme termojenizini artırmaya katkıda bulunduğu ve soğuk iklimli memelilerde daha yaygın olduğu gösterildi.
Sonuç olarak, mitokondri aracılığıyla ısı dağılımının iyileştirilmesine katkıda bulunması beklenen KJD yolundaki değişiklikler, modern EAS popülasyonlarında tespit edildi ve Neandertal ve Denisovan popülasyonları için yalnızca spekülasyon konusu kaldı ancak takip eden fonksiyonel çalışmalar için umut verici lokuslar oluşturdu. Aslında, LEPR rs1137100 çevresindeki varyasyon, EAS'te olumlu seçim tarafından şekillendirilmiş gibi görünürken, yalnızca potansiyel olarak soğuk uyarlanabilir olan LEPR rs1137101, antik türlerde gözlemlendi. Bu, hem modern hem de antik popülasyonlarda KJD yolu aracılığıyla artan termojenizin yakınsayan evrimi veya ilgili antik soğuk uyumlu allellerin modern genomlara entegrasyonunun olası olmadığını gösterir.
Ayrıca, ılıman iklim adaptasyonunun çeşitli modern insan gruplarında farklı şekilde gerçekleştiği gibi görünüyor. Ne yazık ki, kullanılan veri kümesi, çok soğuk ortamlardan popülasyonlar içermiyor, bu nedenle gözlemlenen desenler yalnızca geniş coğrafi alanlarda meydana gelen genel adaptasyonlarla ilişkilendirilebiliyor. Bu, özel yerel adaptasyonların belirlenmesi için potansiyeli kısıtlasa da, EUR gruplarında KJD genleri için seçim işaretlerinin yokluğu, adaptasyonlarının bu genler için sınırlı bir rol içermesine de bağlı olabilir. Bu, diğer yolların (örneğin,