Demir Çağı Britanya'sında kıtasal akın ve yaygın ana yerleşim yeri
Bir toplumun yapısı, evli çiftlerinin ikamet kalıplarından etkilenir. Ortakların çoğunlukla eşin ebeveynleriyle veya yakınında ikamet etmesiyle karakterize matrilokalite, modern etnografik veritabanlarında nispeten nadirdir, patrilokalite ise en yaygın sistemdir. Ayrıca, çoğu Avrupa Neolitik, Bakır ve Tunç Çağı yerleşkesinde, yeterli genomik ve arkeolojik veriye sahip olan yerleşkelerde, patrilokalite ve patrilinealitenin kanıtları bildirilmiştir.
Tarihi dönemin eşiğinde olmalarına rağmen, Britanya Demir Çağı halklarının sosyal yapıları hakkında çok az şey bilinmektedir. Milattan sonraki ilk yüzyıllarda, Ptolemy adanın üzerindeki çeşitli etnik grupların konumlarını Kelt kökenli isimlerle tarif etmiştir (Ek Veri Şekil 1) ve Caesar civitates'i ifade etmiştir. Bu belirsiz terimler genellikle "kabileler" olarak çevrilse de, bu tür grup kimliklerinin karmaşıklığını yeterince anlamıyoruz. İlginç bir şekilde, kaydedilen ilk İngiliz yöneticilerinden ikisi kadın olan Cartimandua ve Boudica'dır, bu da her iki cinsin de en yüksek siyasi statüye ulaşabileceğini göstermektedir. Cartimandua'nın kuzey İngiltere'nin büyük bir kısmını kapsayan Brigantes kabilesinin 30 yıllık yönetimi, kadınların mülk miras edebileceğini, boşanabileceğini ve orduları büyük başarıyla yönetebileceğini göstermektedir. İngiltere'nin doğusunda, Iceni'nin Boudica'sı ünlü olarak üç Roma şehrini yok eden ve imparatorluk hükümetinin otoritesine meydan okuyan bir ayaklanma başlattı. Ayrıca, Julius Caesar, MÖ birinci yüzyılın ortalarında, İngiliz kadınların birden fazla koca alabilir olduğunu belirtmiştir (De Bello Gallico). Ancak, bu tür sosyal açıklamalar, derin bir ataerkil dünyaya dalmış Akdenizli bir izleyici kitlesi için tuhaf görünmüş olan şeye önyargılı olarak kabul edilir.
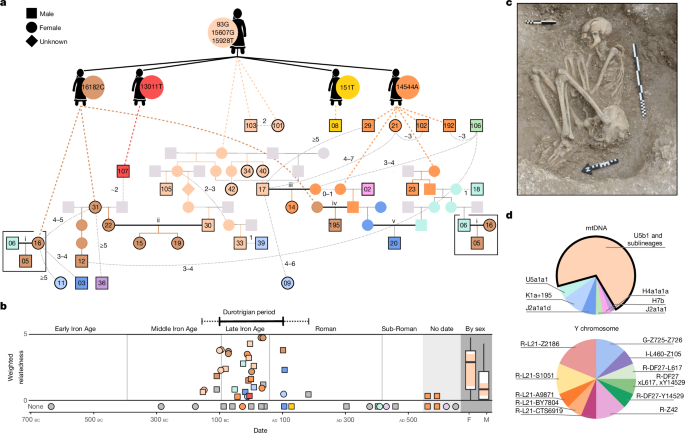
Batı Avrupa Kelt mezarlıklarında bulunan mezar eşyalarının dağılımı, yüksek kadın statüsünü desteklediği şeklinde yorumlanmıştır. Bununla birlikte, İngiliz arkeolojik bulgular sınırlıdır, çünkü Demir Çağı insan kalıntıları nadirdir ve bireyler muhtemelen çoğunlukla yakılmıştır, dışarı çıkarılmış veya bataklıklara gömülmüştür. MS 100 ile MÖ 100 arasında merkezi güney İngiliz kıyılarını işgal eden Durotriges kabilesi, esnek gömülerin resmi mezarlıklarında ölülerini gömmüşlerdi (Şekil 1c ve Ek Bilgi Notu 1). İlginç bir şekilde, bu gömülerde daha fazla sayıda ve çeşitli sayıda prestij eşyasıyla kadınlar daha sıklıkla ilişkilendirilmekte, bu da yüksek statü ve belki de ana odaklı bir topluma işaret etmektedir.
Demir Çağı Britanyalıların genomik varyasyonu araştırılmış, ancak akrabalık ve evlilik ile ilgili toplumsal gelenekleri netleştirebilecek tek bir mezarlıktan sınırlı veriyle incelenmiştir. Genomik tarama, Kelt dillerinin yayılımı konusunda tartışmalara katkıda bulunmuştur (Ek Bilgi Notu 1.6), bu dönemde adaya büyük çaplı bir göçün gerçekleştiğine dair çıkarımlara dayanarak Orta ile Geç Tunç Çağı'nın bu dönem için bir aday pencere olarak kabul edilmesi, Demir Çağı'nda ise önemli bir genetik izolasyonun ardından gelmiştir. Ancak, Britanya'ya gen akışının karakterizasyonu, haplotipik analiz ve bölgesel ayrım yoluyla daha fazla rafine edilmeyi gerektirir. Burada, Dorset'teki Winterborne Kingston (WBK) bölgesindeki diğer mezarlıklardan Durotrigian ve diğer mezarlıklardan 55 genomu ve Maiden Newton ve Langton Herring'den iyi donanımlı iki kadın Durotrigian gömüsünü (Ek Tablo 2 ve Ek Bilgi Notu 1) sıralıyoruz. Bunlar, kadın hatlı soy ile karakterize bir topluluk ortaya koyuyor. Diğer İngiliz Demir Çağı yerleşkelerinden elde edilen verilerle birleştirildiğinde, analizlerimiz matrilokalitenin yaygın olduğunu, coğrafi sınırlarla uyumlu ince ölçekli soyağacı ağlarını ortaya koyduğunu ve güney kıyısındaki Demir Çağı göçünün, hem çağdaş Roma yazıları hem de arkeolojik verilerle uyumlu bir genomik izini gösterdiğini ortaya koymaktadır.
Güney İngiliz kıyısındaki sahil şeridindeki WBK'deki kazılar, MÖ yaklaşık 1000 yılından MS yaklaşık 500 yılına kadar, yani Geç Demir Çağından sonraki Roma sonrası dönem boyunca uzanan, yerleşim için önemli kanıtlar ortaya koymuştur, bunların arasında birkaç küçük Durotrigian türü mezarlık vardır (Şekil 1b ve Ek Bilgi Notu 1). Bölgede bulunan 55 iskelet örneğinden genomik veri elde edildi (Ek Tablo 1), 40'ı genotipe atama ve bireyler arasında özdeş soydan gelen genomik segmentlerin (IBD) güçlü bir şekilde tanımlanması için yeterli bir kapsama ulaştı (>0.3×; Yöntemler ve Ek Bilgi Notu 3). Bu, WBK'nin, yerleşkenin kullanımının Durotrigian dönemi boyunca (MÖ 100 ila MS 100 arasında; Şekil 1b) büyük bir akraba grubunun mezar yeri olduğunu ortaya koydu, 40 kişiden 30'unun yaklaşık yedinci dereceden yakınında en az bir akrabasına sahip olduğunu göstermektedir (Ek Tablo 10; tam ölçütler için Ek Bilgi Notu 4.3'e bakın). Bu akraba grubunun dört düşük kapsama örneği daha, alel eşleştirme analiziyle tespit edildi.
Çarpıcı bir şekilde, genetik olarak belirlenmiş akrabalardan üçte ikiden fazlası (34'ün 24'ü) daha önce antik örneklerde gözlemlenmemiş, modern verilerde sıklığı yalnızca 3 × 10−5 olan nadir bir mitokondriyal haplogroup U5b1 soyu içermektedir (Şekil 1d, Ek Tablo 7 ve Ek Bilgi Notu 2.4). Bu tek matrilinin baskınlığı, sadece iki kardeş çiftin (tüm yetişkinler) gözlemlenmesiyle kardeşlik fazlalığı tarafından bozulmamaktadır (Şekil 1a). Bu haplogroupta, WBK'ye özgü olan dört alt sınıf, ek alt mutasyonlarla ayırt edilmiştir. Mitokondriyal DNA (mtDNA) mutasyon oranının daha hızlı tahminlerinden birini (her nesilde her sitede 4.72 × 10−7 mutasyon) kullanarak, bu düzeyde sınıf içi çeşitliliğin elde edilmesi için soy annelerine en az 420 kadın doğumun gerekli olduğunu tahmin ediyoruz (Ek Bilgi Notu 2.6), bu da bu haplotipin WBK ile uzun vadeli bir ilişki içinde olduğunu göstermektedir. Buna karşılık, Y kromozomu çeşitliliğinin yüksek olduğunu görüyoruz (Şekil 1d ve Ek Bilgi Notu 2.8) ve homosigözlük koşuları (ROH), bu topluluğun dış melezleme ile karakterize edildiğini göstermektedir (Ek Bilgi Notu 5.5). Teori, modelleme ve modern popülasyonların araştırmaları, bu tür kalıpların matrilokal uygulamalar tarafından oluşturulduğunu göstermiştir (yani, erkeklerin dağılımı).
WBK'deki matrilokaliteyi doğrulamak için iki tür simülasyon yaptık (Ek Bilgi Notları 2.10 ve 4.5). İlk olarak, bir popülasyonda demolar arasında erkek ve kadın göç oranlarını modelledik ve sonuçlanan tek ebeveyn haplotip çeşitliliğini (h) tahmin ettik (Yöntemler ve Ek Bilgi Notu 2.2). Bu simülasyonlar, sıfır yakınında bir dış kadın göç oranı ve nesil başına 0.15 ile 1 arasında bir erkek göç oranı göstermiştir. İkinci olarak, üyelerinin sırayla (1) patrilokalite, (2) matrilokalite veya (3) karışık ikamet uygulamaları yaptığı yedi nesilli bir soyağacında otozomal ve X kromozomu akrabalık katsayılarının dağılımını simüle ettik. Yine, gözlemlenen veriler matrilokalite ile uyumludur (Ek Bilgi Notu 4.5). Baskın mtDNA soyunun ilk görülmesi, iki ikinci derece akrabada (MÖ 346 ile 51 arası kalibre edilmiş (cal)) meydana gelmiştir ve MS 31 ile 212 arasında, Britanya Kelt topluluklarının köklü değişimlere uğradığı Roma döneminde son gözlemlenmiştir (Şekil 1b). Buna göre, en son aile üyesi, yeni bir geniş gömülü cenaze töreninin ardından gömüldü (WBK36; MS 82 ile 316 arası cal).
Çekirdek akraba grubu için en tutarlı soyağacını yeniden inşa ettik, bu da WBK'deki matrilokal gelenekleri erkek hareketliliğiyle birleştirmeyi daha da doğruladı (Şekil 1a ve Ek Bilgi Notu 4). İlk dereceden daha büyük tek patrilineal bir ilişki bulduk (WBK02 ve WBK195) ve bu ilişkinin nesiller boyunca matriline kadınlarla çoklu ortaklıklar içerdiğini çıkarıyoruz. Yetişkin bir kadın (WBK31), kızı (WBK22) ve yetişkin torunları (WBK15 ve WBK19) ve ayrıca WBK31'in farklı bir erkek ortak aracılığıyla çıkarılan matrilineal torunu (WBK12) bölgede gömülmüştür. Ayrıca soyağacımızda sıra dışı bir çift ilişki durumu var; IBD segment uzunluğu dağılımından WBK17'nin, ebeveynlerinin evliliğinden kardeşleri WBK34 ve WBK40'ı doğuran evlatlık çocuklarının oğlu olduğu sonucuna varabiliriz (Ek Bilgi Notu 4.8).
Durotrigian dönemine ait bireyleri ele aldığımızda erkeklerin diğer bireylerle önemli ölçüde daha düşük genetik benzerlikler gösterdiğini ve matrilineal bireyler arasında önemli ölçüde fazla temsil edildiklerini görüyoruz (Şekil 1b). Altı birey, hepsi erkek, WBK akraba grubuyla algılanabilir bir genetik bağlantı göstermiyor (yani, baskın matriline'nin üyeleri değil ve akrabaları tanımlanmamış), ancak yine de aile üyeleri olabilir (örneğin, iç göç eden eşler veya evlat edinilen çocuklar). Ölüm anında yetişkin veya ergen olan altı kişiden dördü, tipik Durotrigian tarzında gömüldü, üçü yerel üretim seramik kaplardan oluşan mezar eşyalarına sahipti, bu da toplumun içinde entegre olduklarını gösteriyordu. Genetik olarak akraba bireyleri ele aldığımızda, baskın mitokondriyal haplogroup'a ait olmayan on aile üyesinden sekizinin erkek olduğunu görüyoruz. Bu dış soylu erkekler ve soylu kadınlar arasında iki evlilik yaptığımızı çıkarıyoruz, bunların arasında atalarının çoğunlukla kıta Avrupa'sından geldiği istisnai WBK02 bulunmaktadır (Ek Veri Şekilleri 2 ve 3).
Çiftlerin ortak mezarlarının, erkeklerin sıklıkla matrilineal akrabalarıyla görüşme veya hatta birlikte yaşama ve çoğu zaman eşleriyle değil, onlarla birlikte gömülme eğiliminde olduğu katı matrilineal soyda öncelik verilen bir toplum için tipik olmadığına dikkat ediyoruz.26 Gerçekten de, erkeklerin kayınbiraderleri veya maternal amcaları (avunculate) aracılığıyla miras alma olasılığı nedeniyle erkeklerin eşlerinin ailelerine entegrasyonu, matrilineal sistemleri zorlayabilir.27 Bu nedenle, erkek akrabalarının kontrol edebileceği az mülk olduğunda matrilokalitenin daha istikrarlı olduğu düşünülmektedir. Genellikle kadınlar tarafından bol miktarda ve geniş bir şekilde işlenen ve sahip olunan toprakta ve erkeklerin sıklıkla yokluğunda (örneğin savaş nedeniyle) zenginliğin yoğunlaştığı toplumlara bağlıdır.27,29,30
İlginçtir ki, WBK'de, her iki ortağın da kurucu kadından gelen beş evlilik türü tespit ettik (Şekil 1a), bunların üçünde her iki üye de kadın soyundan gelen doğrudan soyundan geliyor. Bununla birlikte, bu ortakların yavrularında IBD paylaşımı olmaması ve ROH eksikliği ile gösterildiği gibi yakın akrabalığı yoktur (Ek Bilgi Notu 5.5) ve matrilineal çiftler farklı alt sınıflar arasında yer almaktadır. Bu, WBK halkının, yerel bölgedeki ilgili gruplar arasında evlilik düzenlemelerini yönlendirmek için kendi soyağaclarını derinlemesine bildiklerini gösteriyor. Bu modeller, genellikle artan yerel iç evlilik oranlarını (örneğin, yakın köylerden veya aynı köyden bireyler arasındaki evlilik) gösteren modern matrilokal popülasyonlarla tutarlıdır, bu da erkeklerin yerel gruplarında coğrafi yakınlık aracılığıyla etkiyi korumalarına olanak tanır.
WBK topluluğunu bağlama yerleştirmek için, Avrupa'da matrilokal uygulamaların imzası olarak alan ve zaman içinde azaltılmış mitokondriyal çeşitliliği aradık (Ek Bilgi Notu 2.2 ve Ek Tablo 13). Neolitik dönemden Demir Çağı'na kadar uzanan ve birinci derece akrabaları çıkarılan 156 arkeolojik yerleşim yeri dikkate aldık ve hepsi İngiliz Demir Çağı'ndan gelen, çok düşük düzeyde çeşitliliğe sahip altı dışlanmış topluluk gözlemledik (Şekil 2 ve Ek Veri Şekil 1): Worlebury (Somerset), Bottle Knap (Dorset), Gravelly Guy (Oxfordshire), Trethellan Farm ve Tregunnel (Cornwall) ve Pocklington (Yorkshire). Ayrıca, en düşük on bir çeşitlilik tahmininin İngiliz Demir Çağı popülasyonlarından ve bir İngiliz Orta ile Geç Tunç Çağı yerleşim yerinden geldiğini gözlemledik. Buna karşılık, Y kromozomu çeşitliliği yüksektir (Ek Tablo 18 ve Ek Bilgi Notu 2.9) ve ROH desenleri, bunların nispeten büyük dış melezleme toplulukları olduğunu göstermektedir (Ek Bilgi Notu 5.5). Veri kümesindeki ikinci en büyük İngiliz mezarlık örneği olan Pocklington'da, 33 kişiden 28'i üç baskın mtDNA haplogroup'undan birine aittir, bu da WBK'ye benzer şekilde özel mutasyonlarla alt sınıflara ayrılabilir. Burada, ana gömme faaliyeti MÖ 400 ile MÖ 50 arasında gerçekleşti, ancak baskın bir matriline'nin ilk gözlemi, Erken Demir Çağı'nda (I11033; MÖ 717–395 cal; Ek Tablo 12) daha önceki bir tarihte meydana geldi.
Bu sonuçlar, uzun ömürlü matrilokal toplulukların Demir Çağı boyunca ada genelinde yaygın olduğunu ve hatta önceki Tunç Çağı döneminde köken aldığını güçlü bir şekilde göstermektedir. Britanya ve Almanya'daki Zil Kabı ve Erken Tunç Çağı mezarlıklarının analizleri, patrilokalite ve patrilineal soylara vurgu konusunda kanıtlar ortaya koymuştur.12,13,32 Bu durum, Britanya'daki bu dönemdeki daha geniş toplumsal örgütlenmeyi yansıtıyorsa, patrilokal bir toplumun matrilokaliteye geçiş yapma olasılığı oldukça ilginçtir. Bu, etnografik anketlerde nispeten nadir görülen bir durumdur, ancak bunlar insanlık tarihi boyunca çoğu koşulu yansıtmayabilir.4,33
Bir alanda yüksek mitokondriyal çeşitlilik yalnızca ikamet kalıplarını değil, aynı zamanda bireyler arasında genel bir biyolojik akrabalık eksikliğini de gösterebilir; aslında, Demir Çağı Britanya'sında mtDNA çeşitliliği, geliştirilmiş IBD21 kullanılarak tanımlanan akraba çiftlerinin normalleştirilmiş sayısı ile önemli bir (P = 5.85 × 10−7) ters korelasyon göstermektedir (Şekil 2). Bununla birlikte, yüksek biyolojik akrabalık içeren çok sayıda yerleşim yerinin varlığına rağmen, neolitik veya Tunç Çağı Avrupa'sında matrilokal uygulamaların yaygın olmadığını ima eden benzer bir mtDNA çeşitliliğindeki azalma diğer tarih öncesi dönemler için görünmemektedir (Şekil 2). Buna karşılık, İngiliz Demir Çağı popülasyonlarındaki Y kromozomu çeşitliliğini ele aldığımızda, akraba çiftlerinin sayısı ile hiçbir korelasyon tespit edilmemiştir (r = 0,06, P = 0,77; Ek Bilgi Notu 2.9).
İngiltere'nin (İngiltere ve Galler) güneyinden gelen Demir Çağı genomlarında kıta kökenli bileşenlerin artışı tanımlanmış ve adaya Geç Tunç Çağı'nda (yaklaşık MÖ 1000 ila MÖ 875) ve öncesinde büyük çaplı göçler sonucunda olduğu yorumlanmıştır. Bu durum, Erken Avrupa Çiftçisi (EEF) kökenindeki bir artış olarak algılanır (Ek Bilgi Notu 6.2). Verilerimizi dahil ettiğimizde, Erken ve Geç Demir Çağı arasında, merkezi ve doğu İngiliz Kanalı kıyıları boyunca, Durotrigian bölgesini de içeren güney bölgelerinden gelen genomlar tarafından yönlendirilen EEF kökeninde önceden algılanamayan önemli bir artış olduğunu görüyoruz (Şekil 3d ve Ek Tablo 25). Bu bölgeler, yüzyıllar boyunca ölülerin uzaklaştırılmasında, yerleşim mimarisinde ve maddi kültürde paralel gelişmeler sergileyen kanal içi topluluklarla, Tunç Çağı'nın ortalarında benzeri görülmemiş bir kıta etkisinin çekirdeği olarak arkeolojik olarak ortaya çıkmıştır, bu da yüksek nüfus hareketliliğine işaret etmektedir.3 Kanal içi yakın ilişkiler, Britanya'nın büyük bir kısmının daha bölgesel ve ayırt edici bir adalı kültürel iz bırakmaya başladığı Demir Çağı boyunca devam etti.
Genomik veri kümesini "kanal çekirdeği" ve "çevresel" bölgeler olarak böldüğümüzde, Tunç Çağı'nda EEF kökeninin yükselişinin tek bir süreç olmadığını görüyoruz. Aksine, kanal çekirdeği bölgesindeki büyük artış Erken ila Orta Tunç Çağı boyunca meydana gelirken, çevre bölgelerde yüzyıllar süren bir gecikme gözlemleniyor. Örneğin, daha fazla bölgesel bölünme, Erken Tunç Çağı'ndan Erken Demir Çağı'na (yaklaşık MÖ 750 ila MÖ 400) kadar kuzey İngiltere'de EEF kökeninde bir artış göstermiyor (Ek Veri Şekil 7).
Kanal çekirdeği bölgesine özgü kıta gen akışının etkisi, modern ve antik Batı Avrupalılar üzerindeki ana bileşen analizi (PCA) ile (Ek Veri Şekil 2) ve kıta popülasyonlarından haplotip kopyalama kalıplarıyla (ChromoPainter36 kullanılarak) görülebilir (Şekil 3b). Erken Tunç Çağı Britanyalı ve kıta gruplarından gelen katkılara Demir Çağı genomlarının kökenini ayrıştırmak için SOURCEFIND37'yi kullandık ve sonuçlarımızı farklı bir surrogat paneliyle (Yöntemler ve Ek Bilgi Notu 6.3) negatif olmayan en küçük kareler (NNLS) yöntemiyle başka bir yaklaşımla doğruladık. Genel olarak, İngiliz ve Galler Demir Çağı popülasyonuna (MÖ 800 ila MS 50) İngiliz Erken Tunç Çağı'ndan (MÖ 2500 ila MÖ 1500) ortalama %73 katkı sağladığımızı tahmin ediyoruz (SOURCEFIND tarafından tahmin edilen; NNLS tahmini: %75). Bu değer, gen havuzunun %50 uzun süreli yer değiştirmesi oranını çıkardığımız önceki çalışmanın tahmininden daha yüksek olmasına rağmen, rapor edilen İngiliz ve İrlanda'ya özgü R1b-L21 haplogroup Y kromozomlarının dörtte bir oranında seyreltilmesiyle tutarlıdır.
Tunç Çağı sürekliliğinde kanal kıyısı boyunca keskin bir düşüş görülüyor (Şekil 3b ve Ek Veri Şekil 8). Bu, geleneksel olarak Belçik kabileleriyle ilişkilendirilen Hampshire bölgesinde (SOURCEFIND tahmini: %60) merkezlenmiştir ve Caesar'ın Galya'dan göç etmiş olarak belirttiği yerdir. Hem Hampshire hem de komşu Durotrigian bölgesi, Erken ve Geç Demir Çağı arasında EEF kökeninde bağımsız ve önemli artışlar göstermektedir (Ek Veri Şekil 7). Özellikle, Durotrigian bölgesi, Roma etkisinin Galya'ya yayılmasıyla yoğunlaşan kanal içi ağların odak noktalarından biri olan Hengistbury Head'de büyük bir limana sahipti. Analiz için daha az örnek olması nedeniyle, haplotip verileri ince taneli zaman eğilimlerinde daha az çözünürlük sağlar, ancak hepsi kanal çekirdeği bölgesinden gelen ve EEF kökeni tek başına dikkate alındığında ayırt edilemeyen sayısız genetik dışlayıcıyı belirler (Ek Veri Şekil 3; Ek Bilgi Notu 6.3, genetik dışlayıcıların daha fazla tartışmasına bakın). Bu istisnalar arasında, izotop imzası ve gömme töreni temelinde kanal içi göçmenler akımına ait olduğu öne sürülen, Demir Çağı İngiltere'si için bilinen en karmaşık savaşçı mezarlarından biri (kanal kıyısındaki Kuzey Bersted; yaklaşık MÖ 50) yer almaktadır, bu da Julius Caesar'ın Galya'yı fethetmesiyle desteklenmektedir.40
Britanya'nın çeşitli coğrafyası, arkeolojik dönemler boyunca görülen bölgeselliği desteklemektedir. Demir Çağı'nda, nehirler gibi doğal sınırlar tarafından şekillendirilen ince taneli coğrafi genetik yapılar gözlemliyoruz. Çevre bölgeleri (İskoçya, Cornwall, Galler ve kuzey İngiltere dahil) izolasyon izleri göstermektedir. Güney kanal çekirdeği bir istisna oluşturmaktadır, İngiliz Erken Tunç Çağı ile genomik sürekliliğini azaltmaktadır, kanal içi IBD benzerlikleri gösteren yerleşimler, daha büyük nüfus boyutları ve dışsal kökenli bireyler göstermektedir. Bu bölgede, metinsel ve arkeolojik kanıtlarla eşleşen ve en azından sonrasında Roma'nın Galya'ya genişlemesiyle tahrik edilen temas ve değişimin yoğunlaşmasına işaret eden Orta ila Geç Demir Çağı'nda EEF kökeninde bir artış görüyoruz.
Tunç ve Demir Çağları boyunca kanal üzerinden gen akışı, Kelt dillerinin ortaya çıkışı için geniş bir pencere sağlamaktadır. Orta Tunç Çağı'na kadar, kıta kökeninin önemli bileşenleri kanal çekirdeği bölgesinde mevcuttur. Ancak, Demir Çağı'nda EEF kökeninin ikinci bir dalgasının, kanal bölgesinde zaten konuşulan adalı Kelt'in herhangi bir versiyonunu etkilemiş olması muhtemeldir ve güney Britanya (Britonik) ve Galya Kelt dillerinin, İrlanda ve İskoçya'nın Goidelic dillerinde görülmeyen birçok yeniliği paylaştığına dikkat ediyoruz.42 Britanya'nın çoğu bölgesinde Erken Tunç Çağı sürekliliğinin güçlü izleri göz önüne alındığında, bu dönemden sonraki herhangi bir dil girişimi, muhtemelen seçkin bir azınlık tarafından tahrik edilmiş olacaktır.
İngiliz Demir Çağı'nın yaygın matrilokal geleneklerinin kıtadan da getirilmiş olması mümkün, ancak özellikle çevresel popülasyonlarımızda mtDNA çeşitliliği azalmaktadır (Şekil 2 ve Ek Veri Şekil 1). Matrilineal halefiyet, Orta Avrupa'da Hallstatt elitlerinin iki "prens" gömüsü arasında muhtemel bir amca-yeğen ilişkisi keşfedilmesinden yola çıkarak, kıta Kelt toplulukları için daha önce önerilmişti. Matrilineal kurumlar, birleşik ikamet uygulamasına sahip büyük tarım topluluklarında yaygın olan, birleşik soy üzerine kurulu sosyal birimlerin Britanya Demir Çağı'nda da mevcut olabileceğini göstermektedir.4 Ancak, WBK'de erkek eşlerin gömülmesi, bu toplumda matrilineal soy gruplarının varlığının görevlerinde sınırlı olduğunu göstermektedir.26 Matrilokal toplumlarda zayıf bir kayınbiraderlik ilişkisiyle, genellikle anne-kız-kız kardeş ilişkilerinin daha fazla vurgulandığı, kadınların görece daha yüksek statüye ve mülk üzerinde daha fazla kontrole sahip olmaları bekleniyor.27
Hem matrilokalite hem de matrilineallik, kadınların geçim ekonomisindeki katılımını artıran ve babalık kesinliğini azaltan kültürel faktörler tarafından tahmin edilmektedir.28,29,44,45,46 Dış savaş, erkek yokluğu ve çeşitli mekanizmalarla matrilokaliteye geçişi tetikleyen uzun bir süredir teorize edilmektedir.45,47,48, bu hipotez son zamanlarda nicel modelleme yoluyla desteklenmiştir.49 Matrilokalite ayrıca, genellikle sınır savaşlarıyla eşlik eden, yeni bir toprağa göç etme öyküsünü de öngörmektedir.4,45 İngiliz Demir Çağı tartışmasız yüksek toplumsal şiddet dönemi olup, erken dönemdeki höyük kaleleri, silahlar, şiddet ile ilgili yaralar gösteren insan kalıntıları ve Julius Caesar ve Tacitus gibi Roma yazarları tarafından kaydedilen grup çatışmaları örneklerini içerir.50,51,52,53 Önemli olarak, matrilokalite kadınların siyasi ve sosyal güçlenmesini gerektirmekle birlikte, bunlarla güçlü bir şekilde ilişkilendirilmektedir.4,27,54,55,56 ve fethedilen halkların klasik tasvirleri genellikle şüpheyle karşılanırken, bu yazılarımızın Demir Çağı Britanya'sı hakkındaki değerlendirmelerinde bazı gerçekler buluyoruz.
Veri Üretimi
Üç Dorset bölgesindeki üç yerleşimden DNA dizilemesi için 57 gömme örneğini inceledik;2,20,57,58,59 —WBK (n = 55), Langton Herring (n = 1) ve Maiden Newton (n = 1). Çene kemikleri (n = 46), diş kökleri (n = 10) ve tek bir parmak kemiği örnekleri önceliklendirildi. Örnek işleme, Trinity College Dublin'deki antik DNA araştırmasına adanmış temiz oda tesislerinde gerçekleşti. DNA ekstraksiyonu, Ek Tablo 4'te ayrıntılı olarak açıklanan çeşitli protokollere uygun şekilde gerçekleştirildi.60,61,62,63. DNA özütleri, ölüm sonrası deaminasyon hasarlarını azaltmak için USER enzimi ile tedavi edildi ve çift sarmallı kütüphaneler Illumina dizilemesi için oluşturuldu.61,64 Kütüphane bölümleri, örnek özgü indeks primerleri içeren Accuprime Pfx Supermix (Life Technologies) kullanılarak çoğaltıldı (Ek Tablo 5). Çift uçlu veya tek uçlu dizileme, MiSeq, HiSeq 2500 ve NovaSeq 6000 platformlarında gerçekleştirildi (Ek Tablo 5).
Dizi Verisi İşleme
Ayrıştırma için tam P7 indeks eşleşmeleri gerekiyordu; çift uçlu verilerde P5 indeksinde en fazla iki uyumsuzluk kabul edildi. Tek uçlu verilerden adaptörler cutadapt65 ile ve çift uçlu verilerden de AdapterRemoval66 ile çıkarıldı. 11 baz çift örtüşmesine sahip çift uçlu okuma birleştirildi. Tekli okumalar ve kalite ayıklaması gerektiren birleştirilmiş okumalar atıldı. Okuma, varsayılan dışındaki parametreler kullanılarak BWA yazılımı67 ile GRCh37'ye (hs37d5) sahte eşlik eden kopyalarla eşlendi. Okuma, SAMtools68 ile sıralandı, polimeraz zincir reaksiyonu tekrarları Picard Tools v.2.0.1 ile kaldırıldı ve indel'ler GATK yazılımı (v.3.7.0)69 kullanılarak yerel olarak hizalandı. 25'in altında bir eşleme kalitesi ve 34 baz çiftin altında bir okuma uzunluğuna sahip okumalar atıldı. Son olarak, veri, 5' ve 3' uçtaki iki son baz çiftinin Phred kalite puanlarını 2 puana düşürerek "yumuşak biçimde kesilmiş" oldu. Karşılaştırılabilir antik genomik dizi verileri, aynı işleme hattı kullanılarak hizalanmamış FASTQ (mevcut olduğunda) veya BAM (hizalanmış ikili hizalama haritası) dosyalarından indirildi ve yeniden hizalandırıldı (Ek Tablo 12).
Tek Ebeveyn Belirteçleri
Tek ebeveyn belirteci analizinin ayrıntılı bir açıklaması Ek Bilgi Notu 2'de bulunabilir. Kısacası, mitokondriyal haplotip çağrısı için, GRCh37'ye hizalanmış filtrelenmemiş okuma verileri, insan mtDNA'sı için Cambridge Referans Dizisi'ne yeniden hizalandırıldı ve GRCh37 hizalamaları için tanımlanan alt filtrelerden geçirilirdi. Varyantlar BCFtools (v1.10.2)70 kullanılarak çağırıldı ve sonuçlanan VCF (varyant çağrı biçimi) dosyası, Phylotree (Sürüm 17)72'ye dayalı olarak haplogroup'ları atamak için HaploGrep2 (71. referans) girdisine alındı. Kirlilik oranını tahmin etmek için, örnekte bulunan HaploGrep tarafından tanımlanan tek nükleotid varyant sitelerindeki azınlık alellerinin oranını hesapladık (Ek Tablo 7). Arkeolojik alanlar için haplotip çeşitliliği (h), iki rastgele seçilen haplotipin farklı olma olasılığı olarak hesaplandı.73,74 (Ek Tablo 13). Y kromozomu haplotip çağrısı için, okuma işleme hattımızdaki birkaç filtreyi gevşettik: (1) tam P7 indeks eşleşmesi gerekmiyordu; (2) tekli ve kalite ayıklaması gerektiren birleştirilmiş okumaları da içerdik; (3) 20'nin üzerinde bir eşleme kalitesi ve 30'un üzerinde bir okuma uzunluğu için filtreleme uyguladık; ve (4) yumuşak biçimde kesme işlemi gerçekleştirmedik. Y kromozom belirteçleri için Uluslararası Genetik Soybilim Topluluğu (ISOGG) veritabanı (11 Temmuz 2020, sürüm 15.73) ve The Big Tree veritabanı (https://www.ytree.net/) konumundaki konumlarda baz çağrılarını çıkarmak için GATK (v.3.7.0)69'dan gelen Pileup aracını kullandık. 30'un altında bir kaliteye sahip baz çağrıları kaldırıldı. Her erkek örnek için ilgili belirteçlerdeki alel durumu daha sonra değerlendirildi (Ek Tablo 9). Britanya'da Y kromozomu çeşitliliğinin yerleşim yeri tahminlerinde kullanılan haplogroups Ek Tablo 18'de sunulmuştur.
Sahte Haploit Analizi
PCA ve EEF kökeninin nicelenmesinde sahte haploit genotip kullanıldı. İlgili genomlarda tek nükleotid polimorfizmi (SNP) siteleri üzerindeki baz çağrılarını çıkarmak için GATK yazılımından (v.3.7.0)69'dan gelen Pileup aracını kullandık, her site için tek bir baz çağrısı (baz kalitesi >30) seçerek sahte haploit genotip oluşturduk. Batı Avrupa'daki 5,326 modern bireyden oluşan bir veri kümesiyle birleştirilmiş, 1,240k panelinden gelen 534 Demir Çağı bireyi için 1,240k genotip birleştirdi.38 ve yaklaşık 266,000 ortak siteyi kullanarak, antik genomları modern varyasyonun PCA grafiğine smartpca (sürüm 16000) aracılığıyla EIGENSOFT82 kullanarak yansıttık. İngiliz Demir Çağı genomlarında EEF kökenini, önceden tanımlanmış bir yordama göre nicelledik.17 Kısaca, ADMIXTOOLS2 R paketi içinde uygulanan qpAdm aracını kullandık, İngiliz Tunç ve Demir Çağı genomlarını, Batı avcı-toplayıcı, EEF ve bozkır çoban soyundan gelen karışımlar olarak modelledik (Ek Tablolar 12 ve 15). Kaynak ve referans dış gruplar için tam genom dizileri, hedefli SNP yakalama yerine kullanıldı. Kaynak popülasyonlar61,76,79,84,85,86,87, kuzeybatı Avrupa'dan (n = 13) Mezolitik bireyler, Yamnaya çobanları (n = 6) ve Orta ve Güneydoğu Avrupa'dan Erken Neolitik Avrupalılar (n = 9)'dan oluşuyordu. Referans popülasyonlar79,84,88,89,90, Letonya ve Romanya'dan (n = 6) Mezolitik bireyler, Afanasievo çobanları (n = 4), Anadolu Neolitik çiftçilerinden (n = 11) ve Kongo bölgesindeki 10 modern Mbuti bireyden (n = 10) oluşuyordu. PCA ve qpAdm analizleri hakkında daha fazla bilgi Ek Bilgi Notu 6'da verilmektedir.
GLIMPSE Atama
Bu çalışma dahil olmak üzere 2.054 antik bireyden oluşan bir veri kümesinde genotipleme ataması gerçekleştirdik; GLIMPSE yazılımı92 kullanılarak bu çalışma için 42 birey (Ek Tablo 12). Bu hem tüm genom dizileri (>0.1×) hem de hedeflenmiş SNP yakalama (>300.000 çağrı) veri kümelerini kapsıyordu. Atama işle